36. Общий план строения нервной системы рептилий
При масштабном освоении суши мозг рептилий изменился. Однако его морфологические перестройки, на первый взгляд, не были столь принципиальными, как у амфибий. Поверхностный анатомический анализ показывает, что древний «амфибийный» архетип строения центральной нервной системы не подвергся радикальным изменениям (см. рис. III-4; рис. III-5; III-6).
Этот поверхностный взгляд на морфологию нервных центров рептилий может ввести в заблуждение. Только рептилии смогли эффективно освоить сушу, доминировать в воде, подняться в воздух и стать первой доминирующей группой позвоночных. Основой для такой широкой и полиморфной радиации был не только бесконечный пищевой ресурс, но и эффективная система управления организмом — мозг. При появлении рептилий произошёл ряд существенных перестроек нервной системы, которые затем стали базовыми для всех амниот.
По-видимому, эти изменения были схожими, хотя и могли возникать совершенно независимо. Основная трудность состоит в том, что первичный архетип строения нервной системы был замаскирован дальнейшими адаптивными специализациями. Его реконструкция возможна только при выделении ключевых и повторяющихся структурных элементов в организации мозга рептилий. К сожалению, современные рептилии скромно представлены довольно специализированными видами. Это несколько затрудняет поиск древних конструкций мозга, принадлежавших их далёким предкам.
Рассмотрим общий план строения нервной системы рептилий. Их нервная система развита значительно лучше, чем у амфибий и первичноводных позвоночных. Спинной мозг имеет цилиндрическую форму с плечевым и поясничным утолщениями, соответствующими поясам конечностей. Сзади спинной мозг продолжается, постепенно сужаясь, в начало хвостового отдела позвоночника. Головной мозг более широк и массивен, чем у амфибий, особенно у крокодилов и черепах.
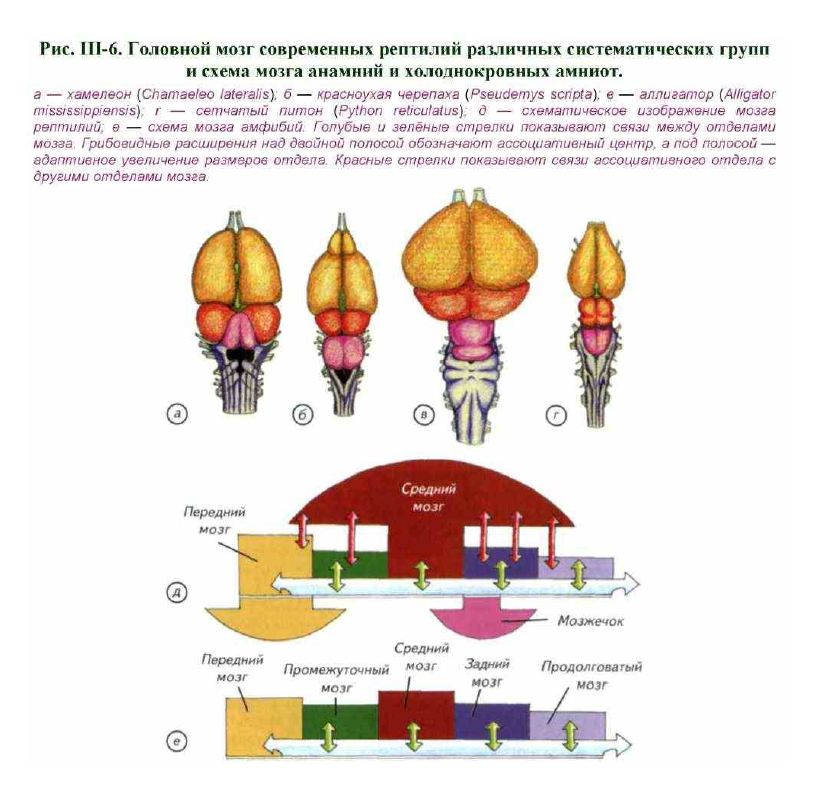
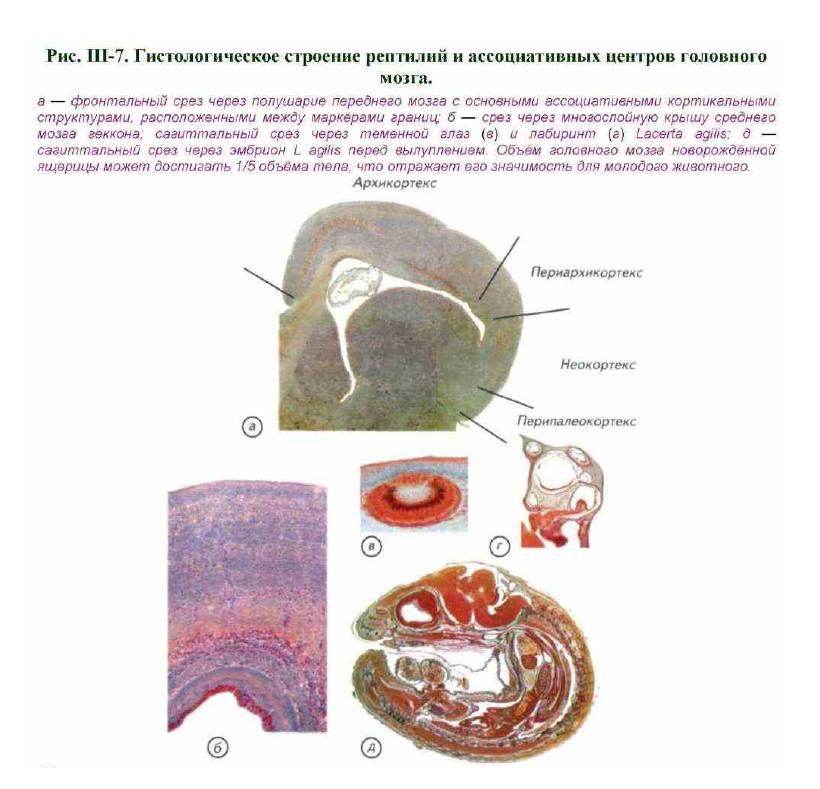
Для рептилий характерны изгибание ствола и стремление крупных полушарий надвинуться на лежащие позади промежуточный и средний мозг (см. рис. III- 6; рис. III-7; III-8). Полушария переднего мозга без резких границ переходят в обонятельные луковицы, от которых начинаются обонятельные нервы.
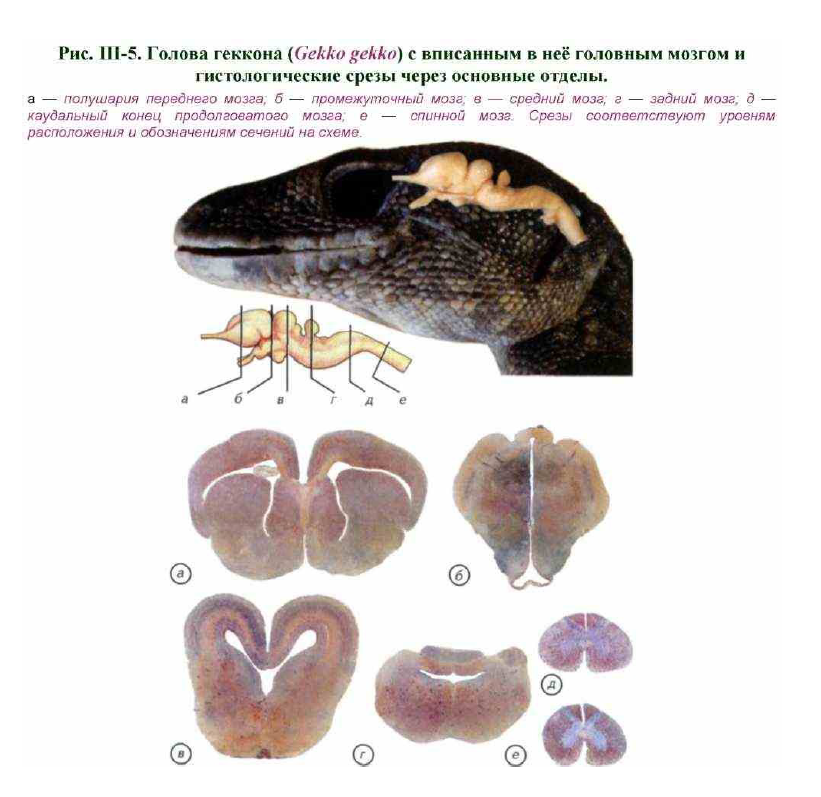
Увеличение размеров полушарий переднего мозга связано с появлением зачатков коры — одного или нескольких слоёв клеток, расположенных под поверхностью мозга (см. рис. III-7, а). В этой зоне переднего мозга осуществляется вторичный анализ информации, поступающей от органов обоняния. Промежуточный мозг почти совсем закрыт сверху передним мозгом, так что на дорсальную поверхность выходит только париетальный (парапинеальный) орган. У многих ящериц этот орган сохраняет строение неинвертированного глаза беспозвоночных(см. рис. III-5-III-8).
Как и у других позвоночных, промежуточный мозг рептилий имеет воронку, переходящую в гипофиз. Средний мозг представлен хорошо развитым двухолмием, которое выполняет функции зрительных центров (см. рис. III-5; III-6). Однако у змей зрительное двухолмие несколько редуцируется и впервые появляются небольшие парные задние бугорки — среднемозговые центры слуха (у рыб и личинок амфибий они обслуживали органы боковой линии).
Двигательные отделы включают в себя задний и продолговатый мозг. Это крупный комплекс мозга с почти закрытым IV желудочком. Он прикрыт сверху мозжечком, который может иметь вид полоски, треугольника или овала, как у крокодилов (см. рис. III- 6).
Черепно-мозговые нервы представлены 12 парами, которые преимущественно обособлены друг от друга. Большинство нервов рептилий не связаны друг с другом и выходят из мозга по отдельности.
Только у змей лицевой нерв иногда тесно связан с узлом тройничного нерва. Обонятельный нерв (I) снабжает своими ветвями обонятельный мешок и якобсонов орган (см. рис. III-8, в). Расположенный рядом терминальный нерв (0) иннервирует этмоидную часть головы. Зрительные нервы (II) образуют в основании черепа перекрёст, называемый хиазмой, при этом у ящериц волокна одного зрительного нерва проходят сквозь щель другого. Лицевой нерв (VII) представляет небольшую, слаборазвитую ветвь. Блуждающий нерв (X) является одним из наиболее длинных черепно-мозговых нервов. Это обусловлено тем, что сердце, желудок и лёгкие сильно удалены от головы.
К блуждающему нерву прилежит добавочный нерв (XI), а подъязычный нерв (XII), начинаясь несколькими корешками от передней части спинного мозга, выходит из черепа через одно или несколько специальных отверстий.
Органы чувств рептилий обладают рядом уникальных особенностей, связанных с ороговением покровов. Несмотря на ороговение, в чешуйках кожи многих рептилий найдены осязательные пятна, а под чешуйками существует развитая сеть осязательных телец с нервными окончаниями. Специализация нервных окончаний, иннервирующих кожу, является прообразом развитых соматических рецепторов млекопитающих (Spearman, Riley, 1980).
Огромное значение как осязательный орган имеет язык. У многих змей и ящериц он постоянно находится в движении. Вкусовыми органами служат луковицеобразные тельца в слизистой стенке языка и нёба. Орган обоняния у рептилий представлен двумя системами: основной обонятельный эпителий носовых мешков) и добавочной (якобсонов орган) (см. рис. III-8).
Носовая полость парная, сообщается с ротовой и глоточной полостью через хоаны. У ящериц и змей часто встречается парная полость, лежащая книзу от носовой и открывающаяся особым отверстием в ротовую полость. На дне полости расположены хемочувствительные карманы — якобсонов орган, в который рептилии вкладывают раздвоенный на конце язык (Halpern, 1987). Слуховой аппарат рептилий состоит из внутреннего и среднего, а иногда и из зачатков наружного уха. Лабиринт более развит, чем у амфибий (см. рис. III-7, г, III-8), а у крокодилов появляются признаки улитки.
У некоторых змей имеются терморецепторы очень высокой чувствительности. Они могут улавливать с расстояния 15 см инфракрасные волны от мыши, температура тела которой выше, чем температура окружающей среды. Так как терморецепторы расположены в лицевых ямках, гремучие змеи могут определять направление на источник теплового излучения. Глаза рептилий обычно несколько крупнее по отношению к голове, чем у амфибий. Глазное яблоко шаровидной формы, а вокруг роговицы у многих ящериц и черепах формируется склеротикальное кольцо из костяных пластинок.
Аккомодация у большинства рептилий достигается при помощи мышцы, расположенной вокруг хрусталика. Эта мышца сжимает хрусталик, делая его толще, и тем самым даёт возможность рассматривать близко расположенные предметы.
Только у змей аккомодация происходит в результате перемещения хрусталика, как у амфибий. В цветных фоторецепторах (колбочках) рептилий и птиц находятся специальные цветные масляные капли, которые сужают кривые спектральной чувствительности рецепторов и уменьшают их взаимное перекрытие. Масляные капли в 2–5 раз увеличивают число различимых цветовых оттенков (Levine, 1985). Многие черепахи обладают хорошим цветовым зрением. В сетчатке одного из исследованных видов Pseudemys scripta) было найдено несколько типов фоторецепторов, отвечающих за восприятие цвета и освещённости: палочки, красные колбочки, красно-зелёные двойные колбочки, одиночные красные и зелёные колбочки. Фоторецепторы работают в различных условиях освещённости фона и объекта, что позволяет черепахам использовать 4 механизма цветовосприятия.
Поведение рептилий построено на врождённых реакциях, которые заметно индивидуализируются в процессе жизни. Этому способствует продолжительность приобретения индивидуального опыта. Крупные черепахи живут до 150–250 лет, змеи и ящерицы — 7-25 лет, а крокодилы — до 70–80 лет. У многих рептилий очень большую роль играет обучение.
Они легко приручаются и быстро осваивают лабиринт или зрительные условные сигналы. Черепахи, принадлежащие к древнейшим анапсидам, могут вторично узнать фигуру, нарисованную сплошной линией, даже если она очерчена пунктиром или повёрнута на небольшой угол. Ящерицы и змеи способны к некоторому прогнозированию развития событий. Это позволяет им преследовать добычу уже после того, как она скрывается из виду. Однако водяные ужи, обученные правильно проходить Т- образный лабиринт, после линьки всё забывают. Аналогично ведёт себя шипохвост, который, привыкнув к новой территории, забывает её после кратковременного охлаждения. Крокодилы проявляют довольно изощрённую заботу о потомстве. Они готовят кучи из земли, листвы и веток, куда откладывают яйца. Пока зародыши развиваются, крокодилы косвенными способами контролируют температуру и охраняют кладку, а затем выкапывают вылупляющихся потомков.
У рептилий известна иерархическая организация компактных групп.
Доминирующей игуане уступают место не только на своей территории, но и на соседних, когда она направляется на охоту в общие нейтральные зоны.
Иерархия у гигантских черепах Галапагосских островов выражается в строго установленном порядке следования друг за другом во время миграций. Групповая организация рептилий является одной из сложных форм общественного поведения, которая была достигнута у холоднокровных животных. Перечисленные особенности современных животных только косвенно отражают зоопсихологические достижения вымерших рептилий. Однако всё разнообразие их поведения базируется на головном мозге общего архетипа, который заслуживает отдельного рассмотрения.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы