29. Формирование мозга позвоночных
Теперь попробуем предположить те события, которые могли привести к развитию описанной выше конструкции головного мозга древних первичноводных позвоночных. Жизнь в мелководной билатерали должна была иметь существенные преимущества для первых хордовых, в противном случае они бы никогда не появились. По-видимому, там небольшие и мягкотелые предки хордовых обнаружили неограниченное количество пищи, множество убежищ и идеальные условия для размножения. Надо уточнить, что перечисленные условия сохранялись достаточно долго, чтобы смогли сформироваться довольно сомнительные конкуренты беспозвоночных — предки хордовых.
Вполне вероятно, что основная конкуренция в этих неглубоких водоёмах происходила между самими примитивными хордовыми без заметного вмешательства со стороны беспозвоночных.
Первые хордовые были явными фильтраторами или собирателями придонных органических осадков. Наиболее эффективные преобразования ротовой полости могли ограничиваться созданием дополнительного ускорения протока воды через ротовую полость и жаберный аппарат. Этого можно было достигнуть как путём развития скелетномышечной системы жаберного аппарата, так и при помощи увеличения подвижности вентральной кожной складки (см. рис. II- 23, в).
Описанные события могли произойти ещё до развития скелетных конструкций в жаберной системе. Существование подобной дочелюстной системы захвата пищи или прокачивания воды могло стать причиной направленной дифференцировки тройничного, лицевого и подъязычного нервов.
Основные эволюционные преобразования в этот период сводились к развитию систем, увеличивающих скорость сбора пищи и минимизирующих затраты на её поиск. Такие проблемы хорошо решаются в билатерали при помощи развития органов вкуса и обоняния. Именно эти события и предполагали формирование обоняния и его аналитического центра — переднего мозга. Параллельно формировался и орган вкуса.
Первоначально это могла быть глоточная часть будущего гипофиза, а затем и специализированные ротовые и кожные вкусовые сосочки. С развитием дистантных обонятельных и вкусовых рецепторов возникла необходимость эффективно двигаться по направлению к источникам пищи. Для этого нужен мало-мальски эффективный механизм плавания. Сегментарная туловищная мускулатура беспозвоночных заложила основы для этих движений, но согласованность её сокращений сложилась вторично на базе сенсомоторных центров заднего и продолговатого мозга.
Любое активное движение в трёхмерной среде приводит к возникновению мозжечка или его аналога. Вполне возможно, что его зачаток возник сразу по двум причинам. Одна была связана с необходимостью плавания в трёхмерной среде, а другая могла быть вызвана возникновением первых электрорецепторных органов.
Мутноватая билатераль вполне располагает к появлению подобных органов чувств, а электрорецепторные органы у беспозвоночных делают такое событие вполне вероятным. Однако описанное хордовое существо не должно было обладать ни одним из перечисленных свойств в совершенстве. Любая самая эффективная, но узкая специализация ведёт в эволюционный тупик.
Далёкий предок хордовых не должен был обладать ни развитой защитой, ни сверхчувствительными органами чувств, ни мощной и дифференцированной мускулатурой для активного движения. Все, кто обзавёлся такими прекрасными приспособлениями для выживания, довольно быстро исчезли.
Неспециализированный предок позвоночных должен был быть своеобразным гадким утёнком с минимумом тактических адаптивных приспособлений. При этом его нервная система должна была обладать рядом свойств, исключающих глубокую специализацию (рис. II-25, а, б): передний мозг не эвертирован, дорсальная часть промежуточного мозга замкнута. Переднее сосудистое сплетение должно было не лежать на мозге, а находиться внутри латеральных и III желудочков. Этому существу надо было обладать не слишком специализированным средним мозгом и мозжечком.
Наиболее вероятный архетип строения мозга такого животного показан на рис. II-25, а. От такого гипотетического предка позвоночных могли взять начало более специализированные формы, дошедшие до нас в виде палеонтологических свидетельств (см. рис. II-25, s). Самым древним известным позвоночным считается Arandaspis. Эти бесчелюстные позвоночные сформировались в ордовике около 500 млн лет назад. Однако
развитые защитные пластины уже покрывали их голову и часть тела. По- видимому, это уже достаточно специализированные существа, возникшие после широкого распространения менее специализированных позвоночных. Дальнейшая эволюция привела к появлению хорошо защищённых бесчелюстных, которые успешно конкурировали с беспозвоночными в мировом океане.
Птераспидоморфные бесчелюстные были уже хорошо защищены мощными панцирями, которые полностью закрывали значительную часть их небольшого тела (см. рис. II-25, в-е). По сути дела, у бесчелюстных был аналог наружного скелета беспозвоночных, они конкурировали с беспозвоночными там, где трудно было ожидать значительного прогресса.
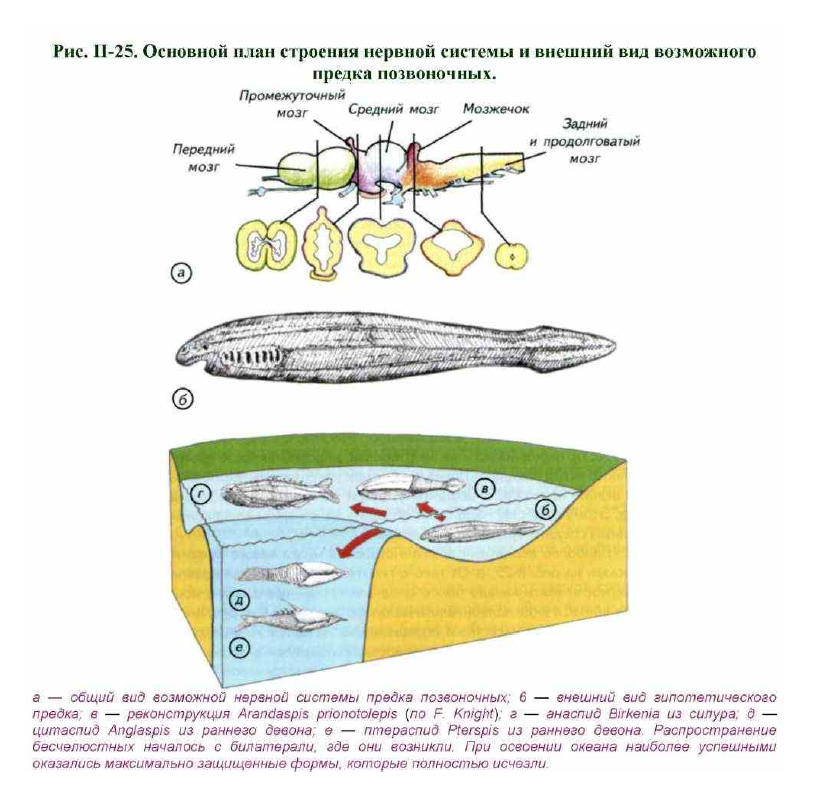
Размеры этих позвоночных были невелики (10–40 см) и сопоставимы с размерами хищных беспозвоночных. Преимущества, даваемые потенциальными возможностями большего размера тела, были ещё не реализованы. Инстинктивный набор поведенческих реакций небольшого мозга древних позвоночных не давал особых адаптивных преимуществ.
Более того, защитный панцирь в передней части тела и плохо защищённый хвост существенно проигрывают полному хитинизированному наружному скелету беспозвоночных, поэтому древние бесчелюстные были неважными конкурентами беспозвоночных в водной среде. Они могли хорошо конкурировать с ними только в довольно специфической среде. Такой средой могли стать мелководные и богатые пищей прибрежные районы
океана. Если плавающий в такой среде предок позвоночных был достаточно подвижен, то он мог конкурировать с менее быстрыми беспозвоночными.
При благоприятных обстоятельствах размер позвоночных мог начать увеличиваться. Впервые были реализованы те возможности увеличения размеров тела, которые даёт позвоночным внутренний скелет. Позвоночное животное могло стать таким большим, что поглощало беспозвоночное
любого размера. Последствия таких эволюционных успехов легко представить. Открылась почти неограниченная пищевая база для эволюционно молодой группы. Это автоматически привело к появлению множества морфологических модификаций общего плана древних позвоночных. Однако, судя по общему плану строения, это произошло уже после появления основных отделов головного и спинного мозга.
Не исключено, что из этой мелководной билатерали одни хордовые шли в более глубоководные части океана, а другие стали приспосабливаться к жизни на границе сред (см. рис. II-25, б-е). Вполне возможно, что именно из этой группы могли появиться предки наземных позвоночных. Это отчасти объясняет отсутствие однозначных палеонтологических свидетельств существования переходных форм. На этот же вывод указывают глубокие различия в морфологии современных первичноводных позвоночных и амфибий.
Происхождение мозга наземных позвоночных Появление наземных позвоночных стало одним из самых занимательных сюжетов в эволюции. Конечно, трудно ожидать, что сравнительный анализ мозга современных амфибий и первичноводных позвоночных даст ответы на все интригующие вопросы этой обширной проблемы. Однако некоторые спорные аспекты происхождения наземных тетрапод можно рассмотреть с неврологических позиций. Этот подход может привести к формально непротиворечивым реконструкциям происхождения нервной системы первых наземных позвоночных.
Основное внимание придётся уделить особенностям строения мозга современных амфибий, поскольку палеонтологические данные в отношении их нервной системы крайне скудны и ненадёжны.
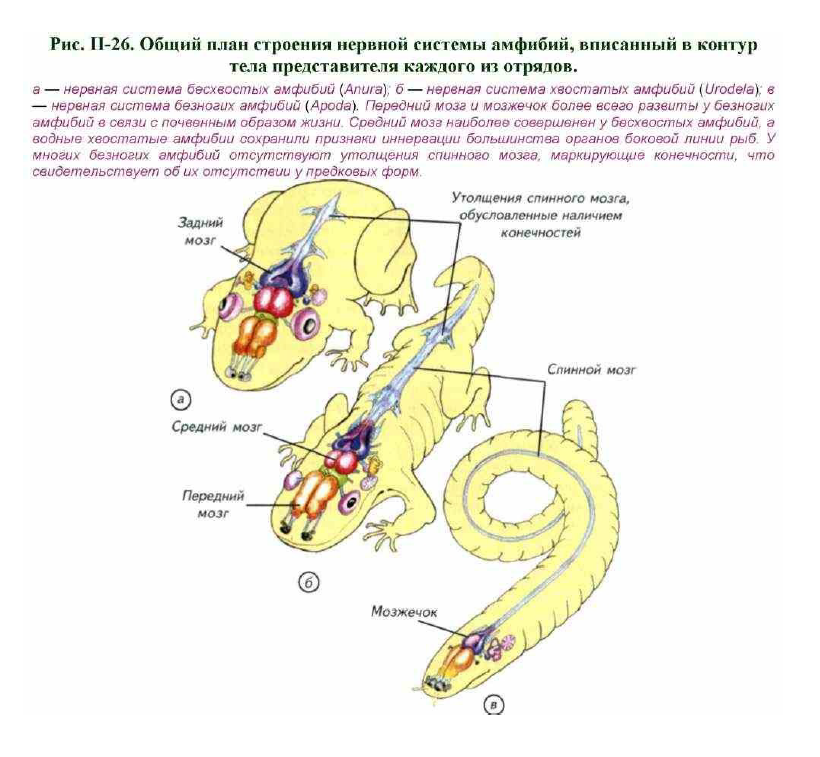
К современным амфибиям относят три относительно самостоятельные группы с довольно туманным прошлым; это хвостатые амфибии (саламандры и тритоны — Urodela), бесхвостые амфибии (лягушки и жабы Anura) и безногие амфибии — червяги (червеобразные земноводные — Apoda) (рис. II-26). Хвостатые и бесхвостые амфибии внешне отличаются от рыб тем, что имеют рычажные конечности вместо парных плавников (см. рис. II-26). В личиночном состоянии почти все амфибии дышат жабрами, которые у ряда видов сохраняются в течение всей жизни. У взрослых животных обычно имеются лёгкие. Личинки амфибий развиваются в водной среде и обладают хрящевым скелетом, тогда как большинство взрослых животных обитает на земле и имеет скелет из костной ткани.
Переход от личиночной стадии к взрослому животному часто связан с выходом на сушу, что приводит к значительным изменениям скелета и внутренних органов. У лягушек исчезает хвост, развиваются лёгкие вместо жабр и формируются конечности. Этот процесс носит название метаморфоза. Некоторые земноводные становятся половозрелыми на личиночной стадии. Это явление лучше всего известно для аксолотля и
носит название неотении. У современных земноводных гладкая кожа, которая легко пересыхает. Амфибии размножаются преимущественно в воде и не имеют тазовых почек, поэтому нуждаются во влажной среде и в большинстве обитают поблизости от водоёмов. Несколько обособленное положение занимают червяги, которые обитают в почве или в водной среде. Они передвигаются в почве при помощи червеобразных движений и отличаются от других амфибий полным отсутствием конечностей и хвоста.
Обитая в почве, они практически лишились зрения, хотя и обладают рудиментарными глазами (см. рис. II-26; рис. II-27-II-29). Среда их обитания наложила глубокий отпечаток на строение органов чувств и головной мозг.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы