8. Заряды мембраны нервных клеток
Однако основные свойства нервной системы обусловлены способностью быстро реагировать на изменение ситуации внутри или вне организма. Скоростные процессы не могут осуществляться по медленным гуморальным законам, они происходят по законам электрохимическим. Нервные клетки способны получать, хранить, перерабатывать и передавать информацию при помощи специальной электрической активности. Они обладают зарядом мембраны — потенциалом покоя и могут его изменять в потенциал действия, который с высокой скоростью распространяется по телу клетки.
В основе потенциала покоя нервных клеток лежит баланс электрохимических и осмотических сил, которые действуют на клеточной границе — мембране. Мембрана клетки полупроницаема. Это означает, что через неё могут проникать далеко не все вещества. Мембрана всегда проницаема для воды, избирательно проницаема для определённых ионов и непроницаема для большинства органических соединений. Молекулы ДНК, РНК, белков и аминокислот находятся внутри клетки и не могут свободно диффундировать через мембрану. В соответствии с законами осмоса вода должна проникать в клетку. Поскольку мембрана клетки непроницаема для органических молекул, осмотическое равновесие достигнуто быть не может. Клетка должна была бы лопнуть. Этого не происходит, поскольку осмотическим силам оказывается постоянное противодействие со стороны сил совершенно другой природы.
Эти силы не осмотические, а электрохимические. Работа осмотических сил уравновешивается работой электрохимических. С одной стороны, это не позволяет клетке лопнуть, а с другой — является источником постоянного заряда мембраны нервной клетки. Внутри клетки находятся молекулы ДНК, РНК, белков, аминокислот и углеводов, которые имеют постоянный заряд. Как правило, этот заряд отрицателен и органические молекулы представляют собой набор внутриклеточных анионов (А). Их заряд уравновешивается внутри клетки положительно заряженными ионами калия (К+). Снаружи клетки основным анионом является хлор (Cl–), а катионом — натрий (Na+). В абстрактной идеальной ситуации
концентрации ионов должны были бы выровняться в результате диффузии через мембрану. Однако внутриклеточные анионы неподвижны, а специальные каналы для всех подвижных ионов обычно закрыты. Более того, специализированные ионные каналы постоянно откачивают избыток натрия и хлора из клетки и закачивают внутрь внеклеточный калий. Это процесс осуществляется с затратой энергии. Она тратится на то, чтобы создать такую величину заряда мембраны, чтобы её хватило для противодействия осмотическим силам, стремящимся разрушить клетку.
В реальной клетке основные проблемы с осмотическими (гидростатическими) силами обусловлены различиями в концентрации ионов калия и натрия по обе стороны мембраны. Некоторую роль в этом
процессе играют ионы натрия и подвижность воды, свободно движущейся через мембрану клетки. Тем не менее основное значение имеет калий, поскольку его концентрационные различия максимальны. Внутри клетки калия примерно в 40 раз больше, а натрия в 9 раз меньше, чем в межклеточном пространстве, поэтому калий стремится уравновесить ситуацию, двигаясь по концентрационному градиенту из клетки, а натрий — в клетку. Поскольку концентрации этих ионов внутри и снаружи клетки известны, можно выразить эти процессы в реальных физических величинах. Работа, которую надо выполнить для предотвращения движения ионов калия из клетки по концентрационному градиенту (А0), будет равна:
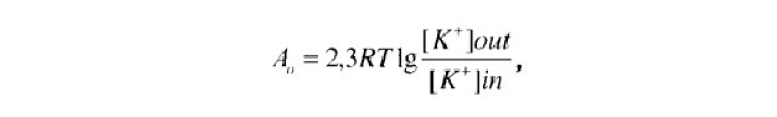
где R — газовая постоянная; Т — абсолютная температура; [К+] — молярная концентрация калия внутри (in) и снаружи клетки (out).
Противодействовать осмотическим силам, стремящимся вывести калий из клетки, могут только электрические силы. Равновесие может быть достигнуто только при равенстве работы осмотических и электрических сил (Аэ):
А0 = А3, Для каждого иона может быть вычислена работа, необходимая для преодоления равновесия электрических сил, возникающих вследствие разделения зарядов по обе стороны мембраны:
A3 = FE,
где F — количество электрических зарядов в моле вещества (постоянная Фарадея); E — выраженная в вольтах разность электрических потенциалов, возникающая в результате разделения зарядов по обе стороны мембраны клетки.
Объединив формулы, легко получить известное уравнение Нернста, или равновесный (диффузионный) потенциал:
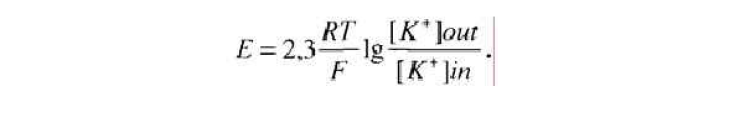
Подставив в формулу равновесия сил реальные значения при комнатной температуре, мы получим реальный заряд мембраны, который называют мембранным потенциалом. Обычно он колеблется в различных клетках от -60 до -90 мВ. При прямых измерениях зарядов мембран нервных клеток получены идентичные результаты. Обычная нервная клетка позвоночного обладает постоянным потенциалом мембраны около -75 мВ. Надо отметить, что подвижны ионы калия, натрия и хлора, поэтому при точных вычислениях мембранных потенциалов надо учитывать другие ионы.
Заряд мембраны и её способность изменять проницаемость для ионов под влиянием различных причин — уникальное эволюционное достижение. Этим свойством обладают многие клетки. Однако нервные клетки используют его для восприятия, передачи и хранения информации. Если при местном воздействии на нервную клетку возникает локальный участок с изменённым зарядом, то сигнал распространяется по мембране нервной клетки. Его обычно называют пассивным, а сам потенциал — градуальным. Это означает, что возникшее локальное возбуждение распространяется по мембране электротонически, что приводит к его постепенному затуханию. Обычно такие сигналы распространяются на небольшие расстояния, хотя у членистоногих известны нейроны, передающие такие сигналы на десятки миллиметров. Градуальные потенциалы образуют светочувствительные клетки сетчатки насекомых и позвоночных, многих периферических рецепторов и даже мотонейронов мышц стенки тела круглых червей.
Потенциал покоя есть у всех клеток на этой планете. Однако в нервной системе он является только отправной точкой для получения, хранения, обработки и передачи сигналов. Информация передаётся в нервных клетках при помощи динамического изменения заряда мембраны клетки.
Это изменение может быть быстрым или медленным. Если оно происходит быстро и изменение заряда мембраны клетки превышает определённый порог, то возникает потенциал действия. Нейроны, формирующие потенциал действия, называют спайковыми. Потенциал действия отличается от градуального потенциала тем, что не затухает по мере движения от места возникновения. Причиной этого является способность мембраны активно распространять местную перезарядку мембраны, если изменение её потенциала достигнет определённого уровня. В обычной ситуации мембрана нервной клетки с потенциалом покоя -75 мВ должна изменить свой заряд до +40 мВ. Такое изменение приводит к формированию незатухающего потенциала действия, который распространяется по мембране клетки. Величину заряда мембраны, с которой начинается распространение сигнала, называют пороговой. Поскольку отростки нервных клеток достаточно хорошо изолированы глиальными клетками, а потенциал распространяется по мембране, его распространение скачкообразное (сальтаторное). В местах, где глиальные клетки плотно прилежат к нейронным мембранам, проведение происходит скачком: от одного перехвата Ранвье до другого.
В перехватах мембрана отростков открыта, что позволяет осуществлять распространение сигнала путём открывания и закрывания калиевых и натриевых ионных каналов.
Принцип передачи сигнала по мембране нервной клетки довольно прост. В его основе лежит высокая плотность ионных каналов в мембране нервных клеток. Количество натриевых каналов может достигать в перехватах Ранвье 12 000 на 1 мкм2. Большое количество ионных каналов в мембранах нервных клеток является основой для распространения потенциалов действия. Натриевые и калиевые каналы расположены довольно плотно, что позволяет формировать локальные встречные потоки ионов при движении сигнала. Потенциал действия образуется в нейроне у аксонного холмика и начинается с открытия натриевых ионных каналов и проникновения натрия внутрь клетки. Поскольку в покое мембрана нейрона имеет заряд внутренней поверхности около -70 мВ, начинается небольшой сдвиг потенциала. Заряд мембраны локально уменьшается и постепенно доходит до нуля, а затем и до +40 мВ.
Изменение знака заряда мембраны приводит к закрыванию натриевых каналов и открыванию калиевых. Поток ионов калия восстанавливает исходный отрицательный заряд. Этот процесс называют кратковременной реверсией потенциала, или потенциалом действия. Возникнув в аксонном холмике, он распространяется по мембране клетки. Волна изменения заряда мембраны двигается с высокой скоростью, а весь процесс реверсии и восстановления занимает миллисекунды. Однако скорости движения потенциалов действия существенно различаются. На это влияют степень миелинизации, диаметр нервных волокон и многие другие факторы.
Скорости проведения потенциалов действия в нервной системе у разных видов животных различаются в десятки раз. Самые высокие скорости известны у аксонов человека и креветок (120–200 м/с), а самые низкие у актиний и медуз (0,1–0,5 м/с). По сути дела процессы передачи сигналов происходят в нервной системе со скоростью, которая отражает динамику и продолжительность жизни организма. Скорость нервных процессов определяет активность животного и внутреннее восприятие времени.
Надо отметить, что передаваемая информация кодируется частотой, последовательностью и продолжительностью активности импульсов, что обеспечивает высокую точность переносимой информации. Эти информационные сигналы различаются не только по принципам кодировки, но и по источникам — типам нервных клеток. Существует морфологическая и физиологическая классификация нейронов. По ведущим функциям нейроны физиологически подразделяются на сенсорные (афферентные), моторные, или двигательные (эфферентные), вставочные (ассоциативные, соединяющие афферентные и эфферентные) и нейросекреторные (гормональные клетки нервной системы). Эти типы не абсолютно жёсткие, а каждая клетка отчасти обладает всеми перечисленными функциями.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы