28. Возникновение отделов головного мозга
Ранний период истории возникновения предков позвоночных, до формирования хорошо структурированного скелета, довольно туманен.
Если допустить, что предковые формы хордовых были мягкотелыми существами размером около 10–15 см, то возникнет существенная проблема как с биотопом, так и с биологическим смыслом появления таких существ. Первым условием возникновения хордовых должна была стать некая очень выгодная среда. В ней должно быть много пищи, ещё не освоенной другими организмами. Эта среда должна давать возможность
эффективно размножаться и защищать от потенциальных хищников.
Вполне возможно, что первые позвоночные возникли в мелководье билатерали. В этих местах крупные морские беспозвоночные были не так опасны, как в воде, а размеры наземных беспозвоночных хищников были намного меньше, чем в водной среде, что позволяло выживать даже мягкотелым предкам позвоночных (Janvier, 1981).
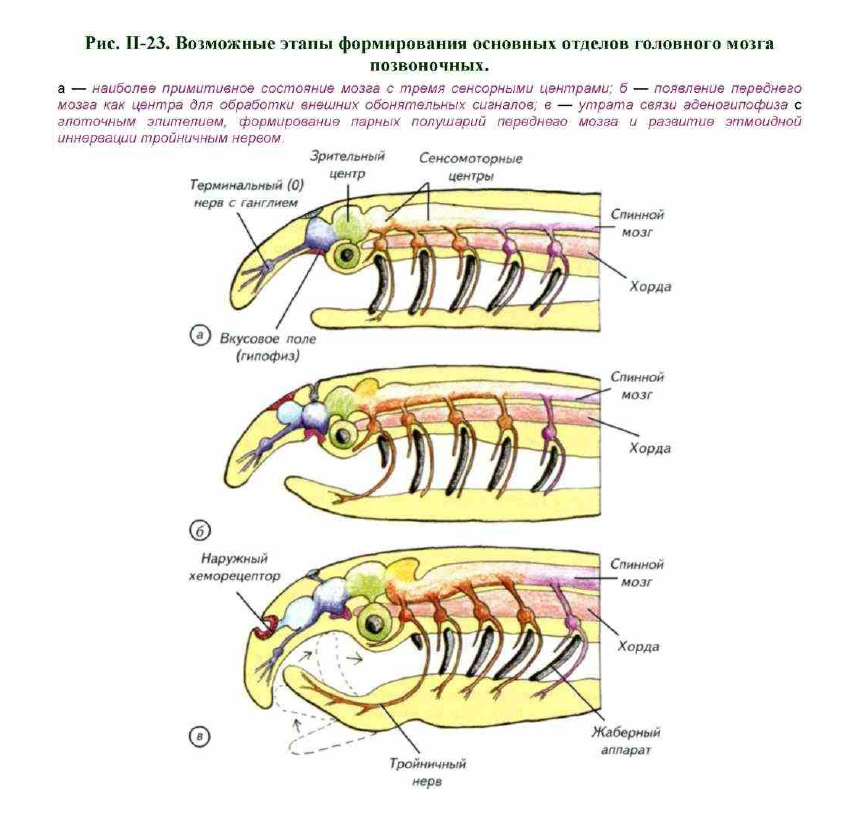
Допуская появление первых хордовых на мелководной билатерали, попробуем представить себе ключевые этапы формирования основных отделов головного мозга. Головной мозг древних хордовых сформировался из 3–4 слившихся ростральных ганглиев нервной цепочки беспозвоночных (см. рис. II-15; II-16). Ганглиозная структура нервной цепочки беспозвоночных предполагает сохранение следов рострокаудальной сегментации, которая отразилась в организации первичных нейральных отделов (рис. II-23, а). В наиболее примитивном состоянии дорсальная нервная трубка состояла в головной части из трёх сенсомоторных центров.
Самой каудальной и самой древней частью были два сенсомоторных ганглия на границе головного и спинного мозга. Они составляли основу координированной эффекторной активности всех моторных нейронов в древней нервной системе. Эта функция была унаследована от беспозвоночных, как и принципы морфологической организации.
Ретикулярно-нейропильная структура продолговатого и заднего мозга современных первичноводных позвоночных очень близка по принципам организации к моторным ганглиозным центрам современных турбеллярий и полихет. Ростральнее моторных центров расположен видоизменённый ганглий, связанный со зрительной системой. Парные глаза также стали наследством беспозвоночных предков.
Маловероятно, что они возникли вторично, хотя инвертированное строение сетчатки и её образование из нервной трубки не исключают и такого варианта развития событий.
Спереди от зрительных центров первоначально располагался ещё один остаток самого рострального ганглия беспозвоночных. Это нейроморфологическое наследство нейрогемального (гормонального) органа. По-видимому, сохранились ганглий и связанный с ним орган гормональной регуляции поведения. Нейрогемальный орган интегрировался в ганглиозную структуру этого участка мозга.
Однако у самого рострального участка нервной трубки возникли и дополнительные функции. В первую очередь это механорецепторный аппарат передней части тела. Эти функции выполнял терминальный нерв (0) с собственным ганглием. Он иннервировал этмоидную зону головы древнего позвоночного так же, как он выполняет эти функции у современных костистых, двоякодышащих и пластиножаберных. Нижняя часть этого прообраза промежуточного мозга соприкасалась с глоточным эпителием, где выполняла функции вкусового рецептора.
В зависимости от состава пищи изменялась гормональная активность нейрогемальных центров и происходило адаптивное изменение как поведения, так и работы пищеварительной системы.
Внутриротовое вкусовое поле было прообразом аденогипофиза, которое впоследствии изменило свои функции. В настоящее время эта гипотеза возникновения и сегментации отделов мозга выглядит наиболее полной, хотя и расходится с предположениями других авторов (Olson, 1986; Keynes, Lumsden, 1990).
Не исключено, что с самого начала эволюции ростральный участок нервной трубки был связан с эктодермой. Примитивные способности эктодермальных клеток к восприятию уровня освещённости позволили сформировать пинеальный глаз. Светочувствительный орган, непосредственно связанный с гормональным центром, давал возможность регулировать суточную гормональную активность. У современных
первичноводных позвоночных эти функции выполняют теменной глаз и нейрогормональный пинеальный комплекс дорсальной части промежуточного мозга (Janvier, 1981; Jollie, 1982).
Судя по всему, внешний дистантный хеморецептор возник несколько позднее (см. рис. II-23, б, в). Он сформировался как ростродорсальное выпячивание части будущего промежуточного мозга в зоне отхождения терминального нерва. По-видимому, он первоначально состоял из непарного утолщения ганглиозного типа и центрального хеморецепторного поля. Однако такая асимметричность сохранялась недолго. Обонятельное
поле разделилось на два самостоятельных органа обоняния с отдельными нервами. Причина таких преобразований хорошо понятна. При непарном органе обоняния надо совершать довольно сложные движения телом, чтобы определить направление на источник запаха. Так поступают современные круглоротые. Увеличение расстояния между двумя сенсорными центрами позволяет точнее и быстрее определять направление на источник запахов без особых затрат на движение.
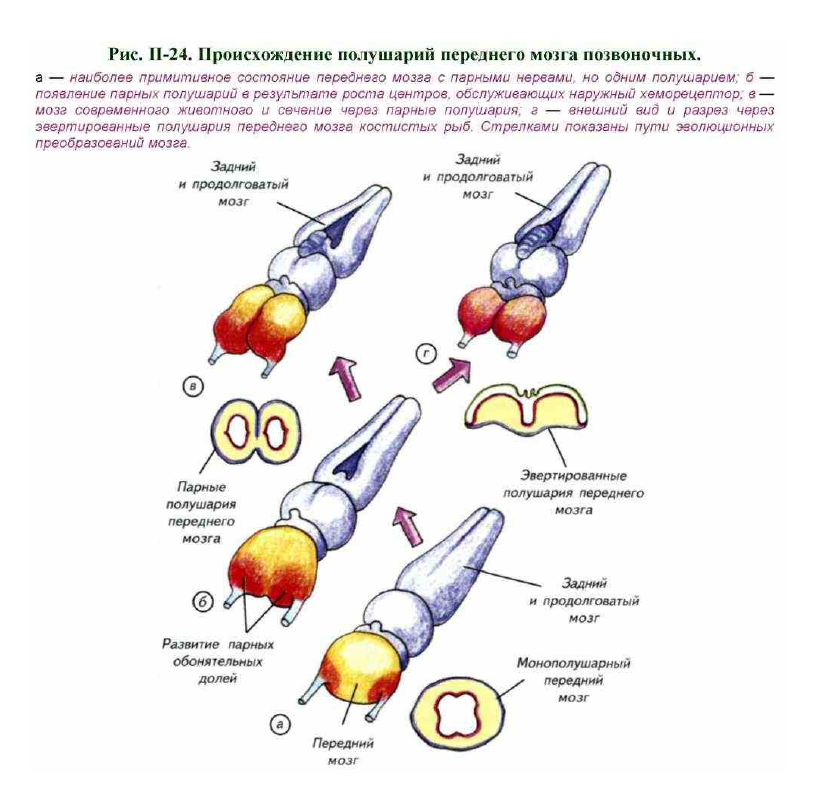
С разделением хеморецепторного поля на два симметричных органа обоняния произошло и другое важное изменение в организации головного мозга — возникли парные полушария (рис. II-24). Можно предположить, что на первом этапе зачаток переднего мозга представлял собой асимметричное утолщение нервной трубки. Затем произошло разделение наружного обонятельного поля и, как следствие, обонятельных нервов (см. рис. II-24, а). Это привело к началу латерального увеличения участков переднего мозга, обрабатывающих обонятельные сигналы.
Параллельно развивались и комиссуральные связи между зачатками обонятельных полушарий. Они были необходимы для сравнения обонятельных сигналов от каждой стороны тела (см. рис. II-24, б). Обонятельные полушария распространялись в ростральном направлении и формировали два слепых кармана — латеральные желудочки. Они и у современных позвоночных соединяются между собой и III желудочком только в каудальной зоне переднего мозга. Эта тенденция увеличения размеров обонятельных центров привела в конечном счете к формированию парных полушарий переднего мозга (см. рис. II-24, в).
Однако наращивание количества нервных клеток в обонятельных центрах мозга первичноводных позвоночных шло различными путями.
Ещё на заре возникновения переднего мозга были реализованы две различные эволюционные стратегии. Одна из них предполагала увеличение размеров переднего мозга путём размножения клеток в прижелудочковом слое с последующей их миграцией в наружные части стенки полушария.
При этом типе развития мозга формируются парные полушария с замкнутой внешней стенкой. Такой вариант организации мы чаще всего встречаем у пластиножаберных (см. рис. II-24, в). У большинства костистых рыб и деление клеток, и их дифференцировка происходят в непосредственной близости от мозговых желудочков.
Следствием такого типа дифференцировки становится выворачивание прижелудочковой поверхности полушарий наружу (см. рис. II-24, г). Эверсия полушарий переднего мозга стала тупиковым вариантом в эволюции нервной системы, если, конечно, считать выход на сушу эволюционно прогрессивным событием. У наземных позвоночных нет эвертированного варианта строения переднего мозга, а его обладатели не смогли покинуть водную среду.
Продолжая реконструировать развитие основных отделов мозга, надо отметить, что с течением времени гипофизарный вкусовой карман утратил своё значение. Его функции стала выполнять часть ротовой полости, которая иннервировалась несколькими ветвями различных жаберных нервов (Mallatt, 1984). Такая множественность иннервации вкусовых клеток сохранилась и у современных позвоночных. Однако рецепторные и секреторные свойства клеток архаичной гипофизарной ямки не исчезли без следа. На её основе сформировался нейрогипофиз, который в сочетании с аденогипофизом стал ключевым органом регуляции желёз внутренней секреции. При переходе к активному плаванию нагрузка на сенсомоторные
отделы возросла, что привело к увеличению размеров задних мозговых отделов, а потом и к образованию IV желудочка.
Эти преобразования уже не были революционными, а возникший архетип мозга сохранился у всех современных позвоночных. Вполне понятно, что проведённая реконструкция возможных этапов эволюции первичноводных позвоночных носит гипотетический характер. Однако сравнительно-морфологические и палеонтологические данные косвенно подтверждают эту точку зрения (Stensio, 1963; Schaeffer, 1981).
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы