43. Нервная система и органы чувств птиц
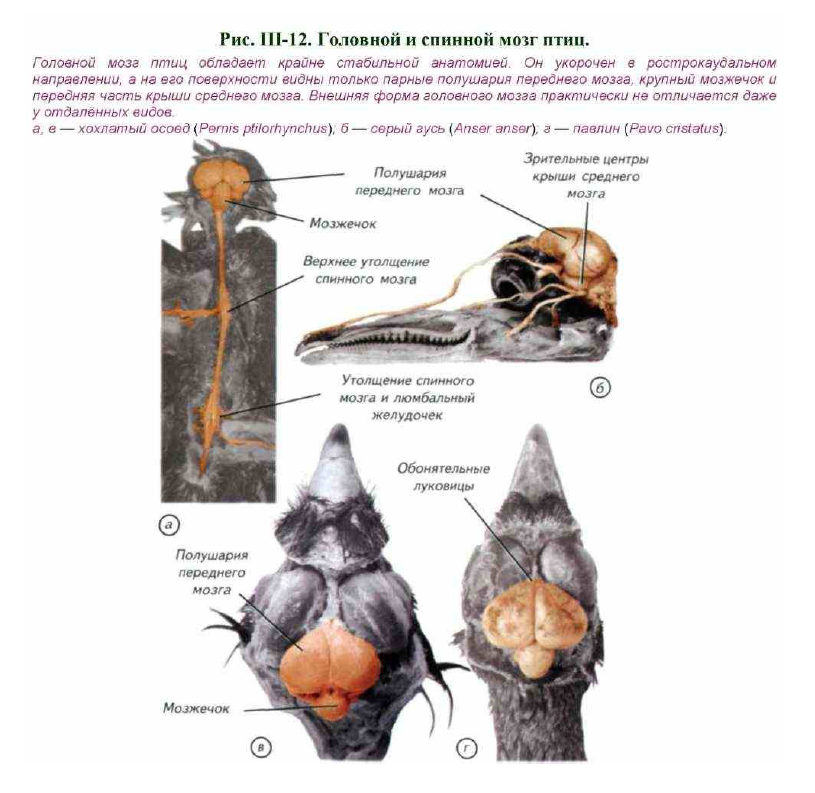
Нервная система птиц состоит из центрального и периферического отделов. Головной мозг птиц крупнее, чем у любых современных представителей рептилий. Он заполняет полость черепа и имеет округлую форму при небольшой длине (см. рис. III-12).
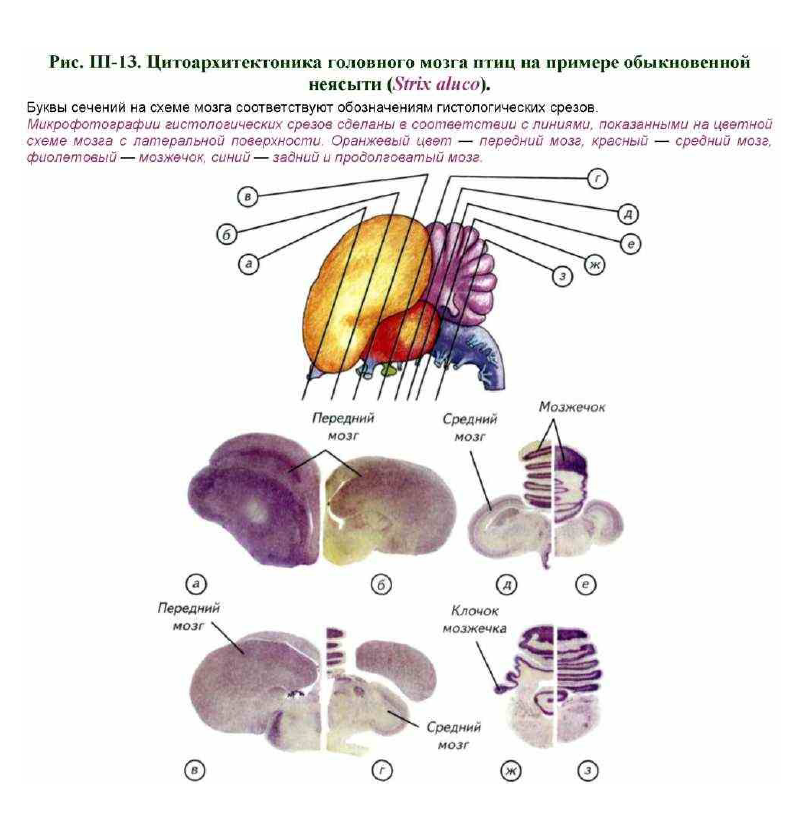
Самый крупный отдел — передний мозг. Он состоит из двух полушарий с гладкой поверхностью или слабо обозначенными продольными углублениями. Эти углубления не являются истинными бороздами, а отражают границы слоёв внутренних ядер (рис. III-13, а, в). Только у попугаев можно заметить небольшое морфологическое обособление височной доли полушарий. Основной объём полушарий занимают подкорковые ядра, кора имеет рудиментарное строение и занимает небольшую часть верхней стенки мозга. Полушария переднего мозга простираются назад до контакта с мозжечком. Следствием разрастания переднего мозга назад, а мозжечка — вперёд является то, что промежуточного мозга снаружи совсем не видно, хотя его можно определить по выросту эпифиза. Эпифиз у птиц развит незначительно, а гипофиз достаточно крупный.
Средний мозг сильно развит, но из-забокового расположения имеет нехарактерную внешнюю морфологию.
Передние выпячивания крыши среднего мозга сдвинуты латерально. Их часто называют зрительными долями (Lobi optici). Мозжечок состоит из массивной средней части (червя), пересекаемой обычно 9 извилинами, и двух небольших боковых долей, которые гомологичны клочку мозжечка млекопитающих. Задний и продолговатый мозг имеет два резких изгиба, обусловленных ориентацией и подвижностью головы птиц.
Гистологическое строение головного мозга птиц существенно отличается от такового у других групп позвоночных. Уже у амфибий можно выявить старый и древний стриатум, септум и в зачаточном виде базальные ядра переднего мозга. У рептилий появляется новый стриатум, который становится доминирующей структурой переднего мозга (см. рис. III-12; III-13, а).
Важно подчеркнуть, что новый стриатум возникает у птиц вопреки развитию зачаточного неокортекса рептилий. Иначе говоря, у рептилий получили значительное развитие структуры стриатума и септума переднего мозга. Они расположены в базальных частях переднего мозга птиц и развиты намного лучше, чем у рептилий (см. рис. III-13, а). Однако рептилии приобрели и зачаточные корковые структуры переднего мозга, которые сформировались в результате развития дополнительной (половой) обонятельной системы. Эти структуры стали выполнять у рептилий функции нового интегративного мозгового центра на базе переднего мозга.
Казалось бы, дальнейшее развитие корковой системы переднего мозга гарантировало бы птицам необходимые поведенческие преимущества. Тем не менее этого не произошло. У птиц корковые структуры, впервые появившиеся в переднем мозге рептилий, носят откровенно рудиментарный характер.
Эволюция корковых структур переднего мозга рептилий полностью остановилась у птиц. Зачатки палео-, архи- и неокортекса практически не играют роли в контроле поведения, поскольку обоняние у птиц развито намного меньше, чем у рептилий. У большинства современных птиц нет развитого обоняния, а вомероназальная система полового обоняния у них вообще отсутствует. По-видимому, в период отделения предков птиц из общего рептилийного ствола обоняние перестало играть для них какую- либо роль. Крупный передний мозг остался функционально невостребован и стал морфологическим субстратом для возникновения ассоциативных центров. Зачаточные корковые структуры не могли играть существенной роли на фоне «освободившихся» от своих функций огромных ядер стриатума и септума.
Невостребованная нейронная система этих структур надолго обеспечила мозг птиц резервами памяти и возможностями развития сложного адаптивного поведения.
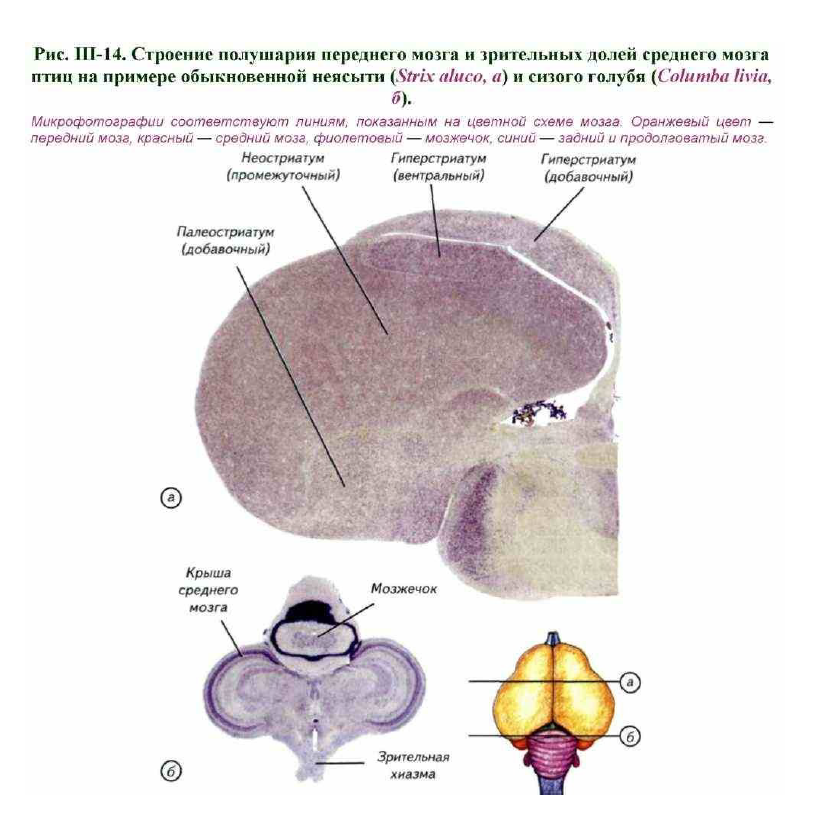
Следы кортикальных структур переднего мозга птиц расположены только в дорсальной зоне полушария, а большую часть мозга занимают стриатум, септум и неостриатум. Функции ассоциативного центра в переднем мозге птиц выполняют чрезвычайно развитые структуры стриатума (рис. III-14, а). Однако историческое название «гиперстриатум» не отражает реального происхождения этого центра птиц. Ранее считалось, что гиперстриатум возник из стриатума и является его гомологом.
Специальные исследования кинетики пролиферации и миграции нейробластов у птиц позволили установить, что гиперстриатум состоит из нейронов различного происхождения.
Они по большей части мигрируют из латеральной (paleopallium) и новой коры (neopallium). Особенностью развития стриатума стало формирование структур ядерного типа, а не стратифицированных корковых образований. У птиц и рептилий разрастание стриатума приводит к экспансии дорсального внутрижелудочкового бугорка, который практически полностью занимает полость латеральных желудочков (см. рис. III-14, а). Гиперстриатум птиц (рептилии имеют его признаки) выполняет функции новой коры. Он представляет собой высший ассоциативный центр, который определяет принятие решений и является основной зоной хранения индивидуального опыта. Через стриатум осуществляются контроль за двигательными функциями и связь с лимбической системой (Northcutt, 1981).
Спинной мозг птиц развит очень хорошо. Он образует большие утолщения в плечевой и поясничной области, откуда отходят нервы передних и задних конечностей (см. рис. III-12, а). В поясничном утолщении верхняя стенка спинного мозга расходится, и центральный канал расширяется в ромбовидный синус, покрытый сверху только озговыми оболочками. Спинномозговые нервы соединяются у птиц корешками ещё в канале позвоночного столба и выходят между дугами или через отверстия слившихся позвонков. В плечевом сплетении обычно участвуют 4 нерва, реже 3, а у страусов — только 1 нерв. Для управления задними конечностями у птиц существует два сплетения: поясничное и седалищное, или крестцовое.
Поясничное сплетение обычно состоит из 3, но у страусов может включать и 5 нервов. Седалищное сплетение у всех птиц состоит из 4 крестцовых нервов.
Черепно-мозговые нервы у птиц представлены 12 парами, которые начинаются по отдельности.
Обонятельный нерв (I) идет от нижней поверхности обонятельной доли, которая спереди и снизу примыкает к полушариям переднего мозга.
Зрительные нервы (II) после хиазмы плавно переходят в средний мозг, а блуждающий нерв, как и у рептилий, идёт из черепа далеко назад, иннервируя сердце, лёгкие, пищевод и желудок. В отличие от рептилий, добавочный нерв (XI) представлен тонкой веточкой блуждающего нерва, а подъязычный (XII) нерв имеет корешки, отходящие как от продолговатого, так и от спинного мозга (Савельев, 2001).
Симпатическая нервная система птиц построена по общему для всех позвоночных принципу. Однако у птиц в шейной части расположен большой симпатический нерв, часто называемый пограничным стволом.
Он лежит в канале, образованном двукорневыми началами поперечных отростков позвонков. Участок этого нерва прилежит к сонным артериям, а далее кзади симпатический нерв находится в канале, образуемом головками и бугорками рёбер, и только в поясничной области он лежит свободно.
По строению органов чувств птицы имеют несколько отличий от остальных животных. Осязание многих птиц (кулики, утки) сосредоточено в челюстном аппарате, где расположены специализированные механорецепторы (тельца Гранди и Хербста). Осязательные тельца расположены в коже у основания крупных перьев и сконцентрированы в восковице, покрывающей основание клюва. Совы обладают особыми осязательными перьями, расположенными вокруг клюва. У дятлов, дроздов, попугаев и глухарей осязательные функции выполняет язык, весьма богатый нервными окончаниями; в нём присутствуют преимущественно различные типы механорецепторов, а не вкусовые сосочки. Птицы пользуются языком как органом осязания. Вкусовые рецепторы у большинства птиц развиты плохо, поскольку птицы обычно заглатывают пищу, не измельчая её в клюве.
Зрение птиц чрезвычайно острое. Глаз способен к аккомодации и обладает рядом морфологических особенностей, позволяющих эффективно ориентироваться, успешно охотиться и издалека определять качество пищи. В сетчатке глаза птиц плотность фоторецепторов намного больше, чем у других позвоночных. В центральной части глаза расположен наклонный гребень, где сосредоточены светочувствительные клетки.
Поскольку оптическая проекция растягивается на наклонной плоскости гребня, птицы видят эту часть изображения увеличенной в 0,25-1,2 раза.
Дополнительным приспособлением, улучшающим цветовосприятие, являются масляные капли в колбочках сетчатки. Они функционируют, как цветовые фильтры, что позволяет различать больше оттенков в световом диапазоне электромагнитных волн. Зрительная система птиц позволяет воспринимать объекты, излучающие в ультрафиолетовом диапазоне, и поляризованный свет. Некоторые перелётные птицы могут непосредственно воспринимать направление электромагнитных полей, что позволяет им ориентироваться в любой точке на поверхности Земли.
У птиц отлично развит слух. Орган слуха состоит из внутреннего, среднего и зачатков наружного уха. Птицы воспринимают звуковые сигналы очень широкого диапазона. В этом отношении они могут намного превосходить многих млекопитающих как по диапазону, так и по чувствительности к слабым звукам. У птиц хорошо развит вестибулярный аппарат. Он состоит из полукружных каналов, отвечающих за рецепцию, связанную с угловым ускорением, и гравитационного рецептора (рецептора линейного ускорения). Все эти компоненты есть и у рептилий.
Однако у птиц появился совершенный мозговой центр анализа вестибулярных и кинестетических сигналов — мозжечок.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы