Нервная система позвоночных
Нервная система позвоночных построена на принципах вероятностного развития, дублирования, избыточности и индивидуальной изменчивости. Это не означает, что в мозге позвоночных нет места генетической детерминации развития или нейрогормональной регуляции. Все эти слагаемые поведения присутствуют. Однако они играют несоизмеримо меньшую роль, чем у беспозвоночных. При развитии нервной системы позвоночных действуют законы формообразования, которые детерминируют первичную региональную экспрессию генов и морфогенез (Савельев, 2001). Одновременно есть и огромная переизбыточность эмбриональных нейробластов. Строгой детерминации развития каждого отдельного нейрона, как у беспозвоночных, нет. Судьба клетки вероятностна и зависит от тех коммуникативных взаимодействий, в которые она вступает во время индивидуального развития и дифференцировки. Достаточно высокая случайность судьбы каждого нейробласта отражается в огромных масштабах гибели клеток нервной системы. При развитии головного мозга млекопитающих нормальная гибель клеток обычно составляет 0,2–0,4 %. К концу метаморфоза бесхвостых амфибий гибнут 85 % всех нейробластов спинного мозга.
Такие потери клеток были бы невосполнимы для беспозвоночных, а у позвоночных эти события составляют нормальный морфогенез.
Вероятностное развитие нервных клеток позвоночных подтверждено в многочисленных экспериментах. Достаточно упомянуть способность нервной системы позвоночных к компенсации искусственно вызванной гибели клеток. Если у эмбриона амфибии на стадии нервной пластинки случайным образом (не в одном месте) разрушить 40 % клеток, то зародыш разовьётся в нормальный организм немного меньшего размера.
Рис. I-15. Гистологическое строение нервной системы позвоночных и беспозвоночных
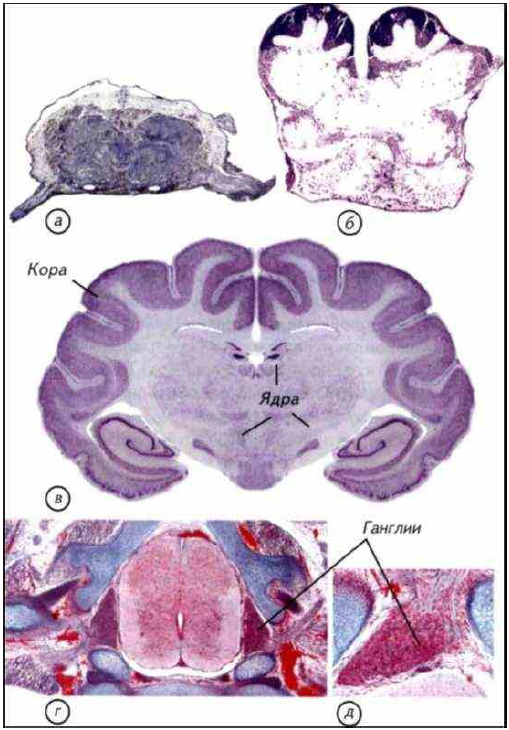
Ганглии нервной системы имеют общий план строения как у позвоночных, так и у беспозвоночных животных. а — подглоточный ганглий речного рака; б — головные ганглии таракана; в — срез через головной мозг хорька; г — спинной мозг домовой мыши; д — ганглии дорсальных рогов спинного мозга мышонка. Срезы а, г, д окрашены по Маллори, б, в — по Нисслю.
Следовательно, нервная система позвоночных уже в эмбриональный период закладывается с «переизбытком» клеток, поскольку их судьба не
столь строго предопределена, как у беспозвоночных. Известные различия в развитии нервной системы позвоночных и беспозвоночных животных
позволяют назвать онтогенетическое развитие позвоночных регуляционным, а беспозвоночных — детерминационным. Это не означает полного отсутствия детерминационных явлений у позвоночных и регуляционных процессов у беспозвоночных. Однако явное преобладание
одного процесса над другим совершенно очевидно. Иначе и не может быть по логике поведения животных этих групп. Жёсткая детерминация развития и поведения свойственна беспозвоночным. Для позвоночных характерны вероятностное развитие и поливариантность индивидуального
поведения. Различные стратегии развития нервной системы позвоночных и беспозвоночных отражаются в основных принципах её морфологической организации. Если нервная система компактизирована из-за размеров и массы тела беспозвоночных, то у позвоночных нет серьёзных физических препятствий для её увеличения. Отсутствие ограничения на размер нервной системы позволяет головному и спинному мозгу позвоночных
достигать крупных размеров и массы 10 кг (Nieuwenhuys, 1998). Однако различия в размерах мозга не исключают сходства гистологической организации у позвоночных и беспозвоночных (см. рис. I-15; рис. I-16).
Нервные клетки в обеих группах животных не имеют принципиальных различий в цитологической организации, хотя некоторые особенности в строении отростков и тел клеток могут быть предметом исследования сравнительной цитологии. В самом простом случае нервные клетки образуют диффузную сеть у животных обеих групп (см. рис. I-16). В ганглиях беспозвоночных и нервной трубке позвоночных клетки и их отростки расположены закономерным образом. К первому типу организации нервных клеток беспозвоночных следует отнести их способность формировать «параллельные ганглии» с однонаправленным вытягиванием тел нейронов и формированием контактов между параллельно расположенными отростками. Такие ганглии мало распространены в нервной системе позвоночных. Второй тип организации нейронов свойствен беспозвоночным и носит название дифференцированного нейропиля, расположенного внутри кольцевого ганглия (см. рис. I-16). Он состоит из тел клеток, расположенных в
корковом слое, и нейропиля, состоящего из переплетённых отростков нейронов. Это наиболее распространённый принцип строения туловищных и головных ганглиев. Третьим типом концентрации нервных элементов в обеих группах является нейропиль с островками тел клеток (см. рис. I-14; I-16).
Рис I-16. Основные тенденции усложнения структурной организации нервной системы.

Синими стрелками обозначен условный этап появления нервныхклеток, красными — основной путь церебрализации беспозвоночных, азелёными — позвоночных животных. В обоих случаях наиболее сложная нервная система организована по ганглиозно-ядерно-корковому типу.Основными отличиями позвоночных являются больший размер мозга,наличие мозговых желудочков и замкнутое кровообращение.
Локальные концентрации тел клеток беспозвоночных сходны с ядрамиголовного и спинного мозга позвоночных. Компактное расположение тел нейронов внутри ганглия позволяет клеткам эффективнее обмениваться информацией. У позвоночных чаще встречаются периферические ганглии, окружённые соединительнотканной оболочкой. Тела клеток в таких ганглиях расположены по всему объёму структуры и не имеют поверхностной локализации, как у беспозвоночных. Нейропиль, состоящий из переплетённых отростков клеток, не выражен. В корковых структурах позвоночных и беспозвоночных расположение тел клеток многослойное (см. рис. I-15; 16). Они обычно разделены пучками волокон, которые позволяют выделять как горизонтальные слои коры, так и вертикальные колонки. Для позвоночных специфично участие разнообразных типов клеток в формировании коры, а у беспозвоночных это довольно редкое явление. Принципиальным морфологическим отличием позвоночных являются мозговые желудочки, которые окружены специализированными эпендимными клетками. Эти клетки сохраняют эмбриональные свойства и могут участвовать в регенерации мозга. У постоянно растущих животных, таких, как неотенические виды хвостатых амфибий, акулы и некоторые рептилии, эпендимные клетки спинного мозга могут пролиферировать и формировать отростки даже у половозрелых особей. У некоторых певчих птиц происходят сезонная пролиферация эпендимных клеток и их миграция в «певческие» центры мозга самцов. Таким образом происходит формирование морфологического субстрата для усложнения песенного репертуара. Сохранение эпендимными клетками способности к пролиферации служит подтверждением регуляционного принципа организации мозга позвоночных. Подобная «резервная избыточность» для беспозвоночных пока не известна.
У позвоночных существует столь же развитая специализация участков головного и спинного мозга, как и у беспозвоночных. Каждый отдел головного мозга представляет собой аналитический центр, обслуживающий один или несколько органов чувств. В зависимости от уровня морфофункционального развития того или иного органа чувств изменяется и морфологическая организация мозгового представительства.
Если один из органов чувств становится доминирующим в поведении животного, то его представительство в головном мозге обычно увеличивается. Как правило, это сопряжено с формированием специализированной ассоциативной «надстройки» в доминирующем отделе. В процессе эволюции это неоднократно приводило к возникновению ассоциативных центров в совершенно разных отделах головного мозга и на различном сенсорном субстрате. Примером может служить среднемозговой центр амфибий, возникший в результате опережающего развития зрения и органов боковой линии. Он имеет стратифицированную структуру, как в грибовидных телах насекомых, и является центром принятия решений и хранения индивидуального опыта.
Многие костистые рыбы, обладающие электрорецепцией, имеют оченьразвитый задний мозг. Именно в нём и его деривате — мозжечке происходит анализ электрорецепторных сигналов из окружающего мира.
Это приводит к появлению стратифицированных структур ассоциативно- аналитического типа уже в заднем мозге. Таким образом, как у беспозвоночных, так и у позвоночных ассоциативные центры являются своеобразной надстройкой над анализаторами и могут располагаться в различных структурных отделах центральной нервной системы. Ум позвоночных нейрогормональные клетки выделены в специализированный
отдел — промежуточный мозг. Он столь же консервативен, как нейрогемальный орган беспозвоночных, и не меняет положения в мозге (Edinger, 1911; Nieuwenhuys, 1998).
Эти данные говорят не только о глубоких различиях, но и о сходстве развития и строения нервной системы позвоночных и беспозвоночных.
Реконструируя пути усложнения морфологической организации мозга, можно предположить такую последовательность событий. На первом этапе исторического развития нервной системы из клеток эктодермального зачатка появились чувствительные элементы (см. рис. I-16).
Специализированные клетки эктодермы обладали одновременно сенсорными и эффекторными функциями. Они рецептировали сигнал, проводили его к эффекторным органам и запускали их реакцию. Эти клетки были связаны между собой и формировали непрерывную сеть,которая не имела выраженных центров (см. рис. I- 12; I-14; I-16).
Такой тип организации нервной системы мы встречаем у кишечнополостных. При появлении более сложных поведенческих задач элементы нервной системы стали объединяться в небольшие скопления. По-видимому, это происходило двумя путями. С одной стороны, формировались параллельные ганглии (см. рис. I-16) с синаптическими контактами между телами клеток. Этот примитивный тип концентрации нервных элементов
отмечен у свободноплавающих кишечнополостных. С другой стороны, появились скопления нейронов с наружным расположением тел клеток и нейропилем из переплетённых отростков внутри ганглия (см. рис. I-16).
Этот тип организации ганглиев оказался достаточно эффективным и сохранился до настоящего времени у большинства беспозвоночных. Такой ганглий обладает рядом преимуществ, которые имеют особое значение для животных с незамкнутой кровеносной системой. Тела его нейронов расположены преимущественно на наружной поверхности, что позволяет поддерживать довольно высокий уровень метаболизма. Через открытую
поверхность тел нейронов происходит снабжение питательными веществами, кислородом и отводятся токсичные продукты жизнедеятельности клеток. Нейропиль, находящийся внутри ганглия, даёт возможность формировать синаптокомплексы, обмениваться сигналами и формировать генерализованный ответ на разнообразные воздействия. По- видимому, из этой формы концентрации нервных элементов возникли головные и туловищные ганглии высших беспозвоночных, ганглии и нервная трубка позвоночных (см. рис. I-15; I-16).
В головных ганглиях беспозвоночных сложились два основных типа гистологических структур: островковые скопления тел клеток и стратифицированные грибовидные тела. Островковые скопления тел клеток беспозвоночных практически идентичны подкорковым и стволовым ядрам позвоночных. Организация грибовидных тел напоминает слоистое расположение клеток в коре млекопитающих. Однако грибовидные тела
беспозвоночных не имеют упорядоченных вертикальных связей между нейронами. Тем не менее стратификация нейронов в грибовидных телах предполагает сходство механизмов обработки информации в ассоциативных центрах позвоночных и беспозвоночных животных.
Вероятно, нервная система позвоночных возникла из ганглиевбеспозвоночных с нейропилем из переплетённых отростков (см. рис. I-16).
Трубчатая нервная система сформировалась в результате выхода отростков нейронов из внутренней полости ганглия. Это событие привело к появлению нейропиля из отростков нервных клеток наружной стороны нервной трубки. Дальнейшее формирование новых нервных центров происходило преимущественно вокруг желудочков, в толще наружного переплетения отростков. В результате возникли центральное серое вещество и окружающие его волокна (белое вещество). Часть клеток выселялась из прижелудочковой зоны и формировала структуры ядерного или стратифицированного типа во внешнем нейропиле (см. рис. I-16). У высших позвоночных центральное серое вещество практически отсутствует, а основные нервные центры мозга представлены сложными ядрами и корковыми структурами различных типов (см. рис. I-15, в). В дальнейшем цефализация позвоночных развивалась по принципу количественного наращивания анатомического представительства анализаторных систем и ассоциативных центров. В отличие от беспозвоночных, нервная система трубчатого типа при замкнутой кровеносной системе может бесконечно увеличиваться в размерах. Это позволило позвоночным достигнуть очень высокого развития умственных способностей.
Подводя итог краткому обзору основных принципов анатомической и гистологической интеграции нервных клеток у беспозвоночных и позвоночных животных, необходимо сделать несколько общих выводов.
Во- первых, нейроны в обеих группах животных имеют сходное строение, но различаются по линейным размерам. Во-вторых, интегративные взаимодействия между нейронами осуществляются в сходных гистологических образованиях: ганглиях, ядрах и стратифицированных структурах. Эти образования встречаются как у беспозвоночных, так и у позвоночных животных.
Основным морфологическим отличием позвоночных являются размер нервной системы, наличие мозговых желудочков и организация кровеносной системы. Эффективная и замкнутая система кровоснабжения позволяет поддерживать высокий уровень метаболизма. Обширная система мозговых желудочков и сосудистое сплетение обеспечивают осмотический и водный баланс головного и спинного мозга. Эти морфологические
особенности позвоночных обеспечивают лабильность, большие адаптационные возможности и размеры нервной системы. Нервная система у беспозвоночных более консервативна, имеет небольшие размеры и не обладает столь широкими адаптивными возможностями, как у позвоночных. Зато она имеет огромный набор генетически детерминированных программ поведения, чётко запрограммированное развитие и эффективную систему гормональной регуляции физиологической активности. Небольшие размеры беспозвоночных позволяют добиваться впечатляющих успехов в конкуренции с другими видами животных. Сравнение организации позвоночных и беспозвоночных отражает различные принципы развития нервной системы, что даёт представление о возможных вариантах её строения и путях эволюции.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы