20. Нервная система с радиальной симметрией
Наиболее простой вариант строения нервной системы мы встречаем у стрекающих (кишечнополостных). Как уже говорилось выше, их нервная система построена по диффузному типу. Клетки образуют пространственную сеть, которая расположена в мезоглее (рис. II-4, а).
Небольшое скопление нервных клеток в окологлоточной области образует подобие распределённого нервного центра. Он интегрирует все несложные реакции тела кишечнополостных и является эволюционным предшественником ганглиозной нервной системы. В окологлоточном нервном кольце формируются параллельные ганглии, описанные в главе (см. рис. I-16). По-видимому, этот тип строения нервной сети был исходным для всех групп животных.
При всей кажущейся простоте диффузный тип нервной системы обеспечивает довольно сложное поведение кишечнополостных. Хорошо известно, что раки-отшельники используют актиний для защиты от хищников. Они выбирают наиболее подходящих особей и пересаживают их себе на раковину. Классическим примером является симбиоз актиний и рака-отшельника. Однако мало известно, что сами актинии также могут выбирать наиболее подходящую поверхность раковины и перемещаться на неё. Иначе говоря, актинии такие же активные, хотя и бессознательные, участники симбиоза, как и раки-отшельники (Холодковский, 1914; Meglitsch, Schram, 1991).
В скромных рамках диффузной нервной системы известно необычайно большое количество вариантов строения. Их всех объединяют радиальная или изоморфная симметрия и общая тенденция к объединению нервных клеток в некие скопления. С момента появления пронейральной сети у губкоподобных организмов началась дифференциальная концентрация нервных элементов. В начале эволюции многоклеточных животных появилось бесконечное разнообразие вариантов строения нервной системы, которые реализовались у кишечнополостных и частично сохранились до нашего времени (см. рис. II-4).
Нервные клетки концентрировались различными способами. Самым простым способом интеграции нервных сетей стало окологлоточное нервное кольцо. Его появление вполне оправдано тем, что оно находилось на границе поступления пищи в организм кишечнополостных. Пища была тем ведущим стимулом, который определял и оценивал успех морфологических изменений нервной системы. Тот, кто мог эффективнее контролировать поступление пищи, увеличивал свой метаболизм и репродуктивные возможности. Самым простым движением, позволяющим проиллюстрировать действие диффузной нервной системы, является реакция на механическое раздражение. Пресноводная гидра (Pelmatohydra oligactis) при малейшем раздражении сжимается в микроскопический комочек. Это происходит за счёт расположенных продольно в эктодерме и поперечно в энтодерме сократимых белков. Кроме генерализованной реакции, кишечнополостные могут дифференциально пользоваться отдельными щупальцами или их группами. Гидры способны
передвигаться, чередуя при переворотах опору на подошву и ротовое отверстие.
Тем не менее диффузная сеть с окологлоточным нервным кольцом была относительно медленно действующим устройством. Измеренная проводимость по нервной сети кишечнополостных составляет не более 5-20 см/с. Этого явно не хватает животным размером более 5 см, поэтому уже у актиний выделились участки нервной сети с высокой скоростью проведения (см. рис. II-4, в). В некоторых случаях она достигает см/с, что делает актиний изощрёнными охотниками за значительно более эволюционно продвинутыми позвоночными. Окологлоточное нервное кольцо было явным достижением, но оно не могло дифференциально управлять всем телом или обеспечить контроль за свободным плаванием.
Среди предков современных одиночных актиний явно были свободноплавающие существа. На это указывает двойная нервная сеть в их теле (см. рис. II-4, б). Одна диффузная сеть расположена под эктодермой в мезоглее и ничем не отличается от таковой у других кишечнополостных (см. рис. II-4, а). Другая нервная сеть лежит в той же мезоглее, но уже около энтодермы. Они связаны между собой только в зоне окологлоточного нервного кольца, которое начинает играть как интегрирующую, так и разделяющую роль. По-видимому, такие двойные сети возникли на заре эволюции нервной системы и были нужны для активного свободного плавания. Животное с автономной «эктодермальной» сетью могло активно двигаться в толще воды.
Сокращение эктодермальных клеток позволяло животному перемещаться, не вовлекая в этот процесс пищеварительную систему
Рис. II-4. Предполагаемые первые этапы (показано стрелками)усложнения строения нервной системы кишечнополостных с радиальной симметрией.
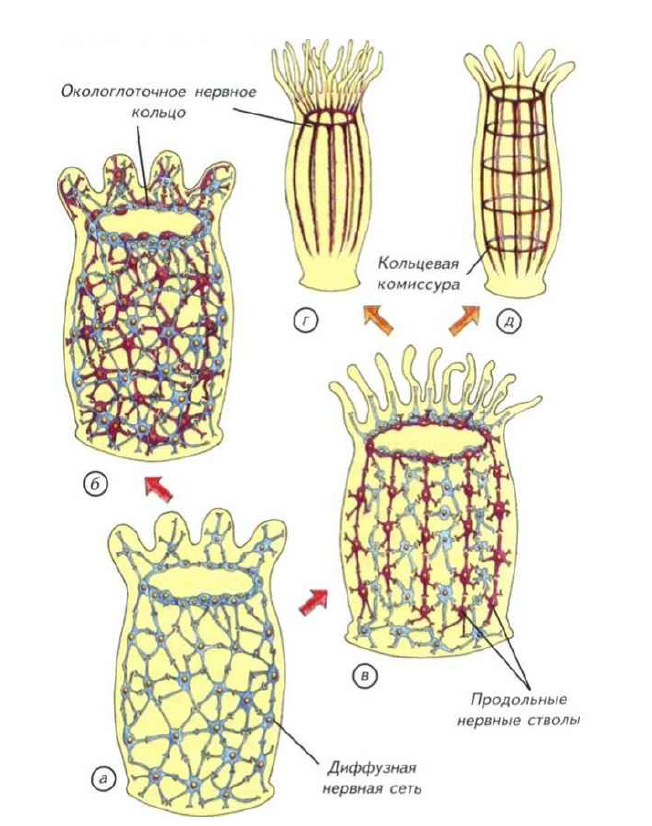
а — однослойная сеть примитивных гидроидов; б — двойная нервная сеть актиний; в — нервная сеть актиний со скоростными проводящими цепями клеток; г — нервная сеть восьмилучевого полипа; д — нервный аппарат радиально-комиссурального типа.
Не исключено, что мезоглея была у этих существ намного толще и рыхлее. Пищеварительная нервная сеть с энтодермальными сократимыми клетками функционировала относительно автономно, перемещая пищевые частицы без эктодермальных сокращений. Такой самодвижущийся пылесос мог быть крайне эффективным при избытке мелких пищевых частиц.
Эволюционные преимущества подвижных фильтраторов хорошо известны, поскольку усатые киты являются самыми крупными животными на планете.
Совершенно иная нервная система у свободноплавающих сцифоидных медуз. Они преимущественно хищники с диффузной нервной системой, которая интегрируется околоротовым круговым скоплением нейронов и несколькими нервными кольцами в зонтике. Эти существа имеют интересные высокоспециализированные участки нервной системы — ропалии. Это небольшие скопления нейронов по краям зонтика. Ропалии
могут содержать статоцисты, или светочувствительные глазки. В статоцистах конкреции различной природы образуют давящий на нейроны «камушек». Он позволяет определять направление на гравитационный центр Земли и ориентировать тело в воде. Глазки измеряют освещённость, а движущиеся волны механически влияют на нервную сеть, что позволяет медузам выбирать направление движения. Подобные нервные образования не смогли стать значимыми центрами для интеграции поведения кишечнополостных, но были первыми специализированными органами чувств. Подобные примитивные рецепторные системы неоднократно возникали в эволюции, что подтверждается разнообразием их структурной
организации при общей убогости рецепторных возможностей.
Можно предположить, что потенциальный предковый вариант строения нервной системы беспозвоночных выглядел как некое кишечнополостное со скоростными тяжами проведения нервных клеток (см. рис. II-4, в). Если допустить эволюционное продолжение концентрации нервных клеток, то из такой морфологической организации с равной вероятностью могла появиться нервная система двух типов строения (см. рис. II-4, г, д). Эти типы различаются только по туловищным комиссурам, которые соединяют продольные нервные стволы.
Окологлоточное нервное кольцо имеет примерно одинаковое строение и интегрирует активность всей нервной сети животного. В хорошо известном плане строения радиально-симметричной нервной системы многих современных кишечнополостных нет поперечных комиссур, соединяющих нервные стволы. Этот тип мог эволюционировать по пути сокращения числа нервных стволов. При этом, по-видимому, возникали самые оригинальные варианты симметрии нервной системы. Примером может служить нервная система нематод (рис. II—5, б). Она представлена 4 параллельными стволами, которые соединяются только окологлоточными нервным кольцом. Других комиссур в глоточной зоне и теле круглых червей нет. Важно подчеркнуть, что 4 нервных ствола нематод расположены симметрично, но вопреки ожиданиям в дорсальном, вентральном и латеральном положении (см. рис. 11—5, б), 4 нервных ствола иннервируют треугольный рот и не имеют отростков, проникающих в мышечные клетки. Наоборот, мышечные клетки образуют отростки, которые оканчиваются на дорсальном и вентральном нервных стволах, идущих вдоль тела. Каждая мышечная клетка имеет несколько таких отростков, а сократимые белки локализованы в дистальном участке цитоплазмы. По этим отросткам проходит нервный сигнал, который и заставляет сокращаться группы мышечных клеток.
Вполне вероятно, что у нематод сохранился древнейший механизм «информирования» клеток организма co стороны нервной системы.
Мышечные клетки сами заботятся об источнике информации, пригодном для повышения метаболизма. Такой тип нервно-мышечных связей крайне примитивен и претендует на эволюционную первичность, что косвенно подтверждает высказанную ранее гипотезу происхождения нервных клеток. Нематоды многочисленны, но не разнообразны по строению органов чувств. В основном это внешние и внутренние механорецепторы,
хеморецепторы (чувствительные ямки, сосочки) и простые глазки.
Механорецепторы специализированы на мужские сенсорные органы и спикулы, головные и соматические щетинки. Однако это пример крайней специализации, который показывает, что наиболее эволюционно перспективным был «комиссуральный» путь (см. рис. II-5, в).
Комиссуры, посегментно связывающие продольные нервные стволы, дают существенные преимущества в дифференциальной активности отдельных участков тела. Вполне возможно, что комиссуральные нервные стволы сформировались ещё на уровне гипотетических кишечнополостных с радиальной симметрией. Множественные нервные стволы таких животных могли иметь комиссуры, которые создавали развитую пространственную нервную сеть. Неподвижным особям вполне достаточно бескомиссурного варианта (см. рис. II-5, а), поэтому комиссуры свидетельствуют скорее о подвижном образе жизни. Сегментированная сеть носила вполне практический характер и использовалась для перистальтического движения. Животное двигалось в результате распространения кольцевых перистальтических волн по телу назад относительно движения. Дифференциальное управление такими полостями и окружающими их мышцами возможно только при наличии повторяющихся нейральных сегментов. В таком сегменте должен быть
автономный центр, управляющий мышцами, — ганглий. У радиально- симметричного животного их может быть несколько, у билатерально-симметричного — 2 или 4.
Такие ганглии расположены в узлах пересечения нервных стволов и поперечных комиссур.
Пересечения постепенно трансформируются в контактные узлы, а затем и в полноценные ганглии. Появление дополнительных периферических центров позволяет им принять на себя часть забот об управлении телом животного. Сегментарные комиссуры с ганглиями являются основным условием возникновения внутри тела специализированных полостей и целома. Без развитой посегментной иннервации септально-целомические
конструкции не имели бы биологического смысла. Их использование для перистальтических движений было бы невозможно. Развитая иннервация позволяет деформировать их в широких диапазонах и развивать большие усилия при различных способах перистальтического движения.
Следовательно, комиссуры и узловые ганглии создали у радиально- симметричного животного все предпосылки для возникновения сегментарности и билатеральной симметрии.
Радиально-симметричное животное, похожее на трубу с пробегающимипо ней волнами, не самый лучший пловец. Этот тип движения очень эффективен в почве, но в воде преимущество получают животные с меньшим числом осей симметрии. Плоскотелые животные с волнообразными движениями тела двигаются быстрее, а их энергетические затраты ниже. Это касается как придонной зоны, так и толщи воды. Замена радиальной симметрии на билатеральную была делом очень небольшого времени. По-видимому, уменьшение числа продольных нервных стволов происходило путём их слияния. Стволы сближались и сливались, как это происходит при метаморфозе насекомых. Мы не знаем, из какой радиальной системы складывалась билатеральная нервная система, но маловероятно, что в ней было нечётное число нервных стволов. В конечном итоге слияние продольных стволов привело к возникновению билатерально-организованной нервной системы. Вероятнее всего, билатеральность сложилась в придонном слое. Древнее
свободноплавающее существо перешло к придонному образу жизни.
Успешно передвигаться внутри придонного слоя могла и радиально- симметричная трубка. Однако более эффективно плавать или ползать по поверхности может билатерально-симметричное существо. Такой тип организации нервной системы широко распространён и среди современных свободноживущих плоских червей — турбеллярий. Встречаются варианты строения с 4 и 2 параллельными нервными стволами (см. рис. II-5, г, е).
Рис. II-5. Общий вид и сечения основных вариантов строения нервной системы кишечнополостных и червей.
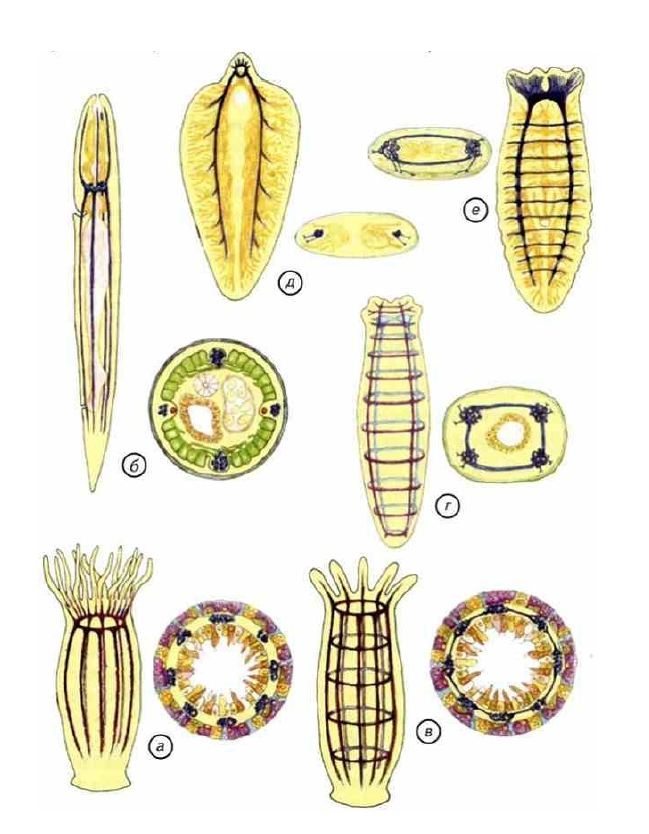
а — нервная сеть восьмилучевого полипа; б — нервная сеть нематод; в — нервный аппарат радиально-комиссурального типа; г, е — нервная система тубеллярий; д — нервная система печеночного сосальщика. Синим цветом на сечениях обозначены нервные стволы.
Судя по всему, примерно с такого набора нервных устройств началась эволюция симметричных нейральных конструкций. Надо отметить, что все достоинства описанной нервной системы не могут «противостоять» обильной пище и возможности бесконтрольно размножаться. Идеальные условия обитания или гиперспециализация разрушают любую нервную систему. В связи с этим у паразитических червей часто отсутствуют поперечные комиссуры (см. рис. II-5, д), а иногда и вся нервная система.
Тем не менее некоторые свидетельства существования множественных продольных нервных стволов можно встретить даже у трематод. Их нервная система состоит из трёх пар неравноценных нервных тяжей, проходящих вдоль всего тела, похоже устроена нервная система паразитической аскариды. У неё два главных нервных тяжа выполняют основные функции, хотя и не имеют комиссур. Кроме этого, существуют 4 более тонких нервных ствола с второстепенными функциями. Эти примеры крайней специализации показывают, что на первых этапах эволюции различные формы радиальной симметрии были заменены билатеральной
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы