17. Признаки мышления
1. Любые генетически детерминированные формы поведения не могут считаться мышлением. Такая детерминация может иметь как прямой генетический характер, так и косвенные формы. Генетически детерминированные формы поведения могут реализовываться через гормональную активность эндокринных систем или регулироваться внешними факторами.
2. Мышление всегда индивидуально. Действие животного и человека, являющееся плодом мышления, не может быть абсолютно идентичным у двух особей. В основе расхождений лежат индивидуальность и изменчивость морфологической организации головного мозга. На разном структурном субстрате не возникает идентичных решений.
3. Биологической целью мышления является достижение поведенческой исключительности конкретной особи. Этим благоприобретённым способом расширяется количество вариантов поиска пищи, размножения и доминирования. Результаты поиска нестандартных решений могут равновероятностно приводить как к положительному, так и к отрицательному (понижающему достижение перечисленных целей) итогу, поэтому эффективность мышления заведомо ниже, чем реализация врождённых форм поведения.
4. Поведение, ставшее плодом мышления, передаётся следующемупоколению с помощью непосредственного научения. Распространение вновь «выдуманной» формы поведения происходит от особи к особи или от родителей к детям, что создаёт основы для социального наследования информации.
5. Мышление — только потенциальная способность мозга. Способность построена на индивидуальных особенностях архитектоники нервной системы. При исключительно благоприятных условиях способность к мышлению может никогда не реализоваться за время жизни конкретной особи. Для реализации мышления необходимо полностью исчерпать все врождённые формы поведения. В целом мышление — самое невыгодное занятие для мозга. Трудно придумать более порочный способ тратить драгоценную энергию нейронов. Вся история эволюции нервной системы построена на стремлении к экономии расходуемой энергии, а не на её трате. Мышление не является исключением из правила.
На «разумной деятельности» всегда принято экономить. Любая интеллектуальная нагрузка крайне затратна для организма, поэтому в эволюции мозга сложились изощрённые способы сохранения энергии.
Для большинства беспозвоночных и позвоночных животных выход был найден через генетическую детерминацию различных форм поведения. Это прекрасный способ избежать излишних затрат на интеллектуальную деятельность. Совершенно ясно, что врождённые формы контроля
поведения предшествовали мало-мальски осмысленному поведению (Шимкевич, 1907). Тем не менее даже детерминированное поведение нуждается в адаптации к внешней среде. Нельзя допустить, чтобы спаривающийся кот безрассудно увлёкся пробегающей мышкой. Эти события надо как-то развести во времени. Стали развиваться системы получения адаптирующей информации — внешние и внутренние органы чувств. Развитие зрения, обоняния, вкуса, слуха, барорецепторов,соматической чувствительности, терморецепторов, осморецепторов и многих других систем привело к увеличению размеров их аналитической части — мозга.
Увеличение массы мозга неизбежно вызвало к жизни генерализованный, но достаточно адаптивный способ управления организмом. Этим механизмом стала гормональная регуляция основных форм поведенческой активности. Развитость органов чувств позволяла точно выбирать нужную врождённую программу, а гормональная регуляция становилась и пусковым стимулом, и системой контроля за достижением адекватного результата. В качестве простейшего случаяможно рассмотреть пример с половым поведением. В период гона самец лося проявляет невероятную активность в конкуренции за самку и стремлении спариться, но после успешных спариваний полностью меняет стратегию поведения и переключается на поиск пищи. Изменение концентрации половых гормонов вполне заменяет мышление. Сочетание наследуемых программ поведения, развитых органов чувств и гормонального контроля реализации форм поведения приводит к эффективному адаптивному поведению. Оно часто вполне достаточно даже для животных с крупным мозгом. По сути дела, если такая форма регуляции поведения оказывается успешной, то никакого «затратного мышления» ожидать от животного или человека не приходится.
Тем не менее сравнивать информацию от различных органов чувств в головном мозге, в какой-то его области, необходимо. В связи с этим ещё на заре эволюции центральной нервной системы возникли специальные центры. В них сравнивалась информация от внутренних и внешних рецепторов, эффекторных органов и принимались «решения», крайне далёкие от того, что мы понимаем под мышлением. Однако даже простой выбор инстинктивного поведения требует некоего специализированного участка мозга, где бы могло осуществляться относительно «беспристрастное» сравнение всех сигналов из внешней среды и организма. Чем объективнее такое сравнение, тем выше вероятность совершить адекватный поступок, увеличивающий адаптивность организма.
Правильный выбор будет правом на жизнь. Эти жёсткие требования и привели к формированию своеобразной надстройки над рецепторными и эффекторными системами — ассоциативных зон мозга (рис. I-22). Ассоциативные центры далеко не одинаковы в различных систематических группах позвоночных. Как отмечалось выше, они неоднократно возникали в процессе эволюции на основе самых разнообразных отделов головного мозга. В далёком прошлом у первичноводных животных ярко выраженные ассоциативные центры отсутствовали. Их свойства были распределены между различными отделами мозга (см. рис. I-22). Каждый ассоциативный центр базируется на определённом рецепторном или моторном центре. Так, у хрящевых рыб практически равными ассоциативными возможностями обладает обонятельный передний мозг, зрительный средний и моторный задний.
Связи между этими ассоциативными центрами весьма слабы, и только очень заметное преобладание одного из них влияет на выбор стратегии поведения. В этом случае он на некоторое время становится ведущим ассоциативным центром. Через некоторое время другой орган чувств окажется ведущим и роль главного ассоциативного центра возьмёт на себя другой отдел.
Это простой, но самый эффективный способ принятия решений при очевидном недостатке нейронов, не обременённых прямым обслуживанием органов чувств. Такой вариант организации ассоциативных центров неоднократно реализовывался в процессе эволюции на различном нейрональном субстрате. У рептилий ассоциативные центры расположены в среднем и переднем мозге, основную роль играет дорсальная часть
среднего мозга.
Рис. I-22. Головной мозг основных представителей позвоночных с латеральной поверхности.
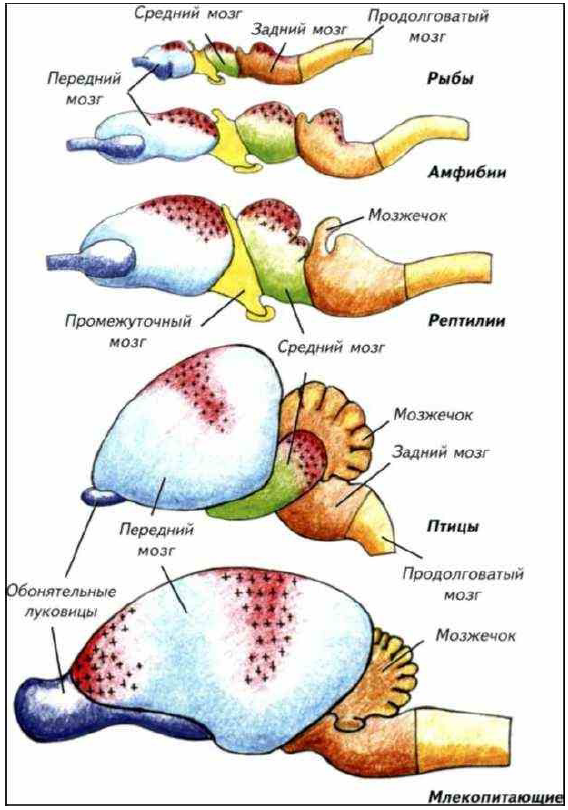
Основные отделы мозга обозначены одинаковым цветом. Красный цвет и крестообразная штриховка показывают расположение ассоциативных зон мозга.
У птиц преобладают ассоциативные центры переднего мозга, хотя роль зрительных центров среднего мозга остаётся значительной. Только у млекопитающих ассоциативные центры сосредоточены в развитом неокортексе переднего мозга. Это стало возможно благодаря увеличению размеров мозга в целом и переднего мозга в частности. Избыток клеток в анализаторных и ассоциативных зонах дал огромные преимущества
животным и существенно усложнил мышление.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы