32. Появление конечностей
На первый взгляд происхождение конечностей наземных позвоночных не вызывает каких-либо логических проблем. Их появление вторично по отношению к миграции предков наземных позвоночных на мелководье и формированию эффективного воздушного дыхания. Вывод основывается на том, что у большинства рыб с воздушным дыханием конечностей нет или они используются частично. По-видимому, использование плавников в качестве конечностей началось только тогда, когда изгибание тела в горизонтальной плоскости оказалось недостаточно эффективным для движения. Представить себе такую ситуацию довольно трудно.
Ископаемые Sarcopterygii (мясистолопастные) обладали относительно развитой мускулатурой плавников, которая позволяла передвигаться по дну или отталкиваться от субстрата. Однако это не основной, а только вспомогательный способ движения. Учитывая массу тела и размеры плавников рипидистий, можно сказать, что их применение для активного передвижения сопряжено с большими трудностями. Даже если допустить,
что они были точкой опоры для более эффективного изгибания и переноса веса тела, причина их трансформации в активную конечность всё равно остаётся неясной.
Возникает парадоксальная ситуация. Зачатки конечностей могли возникнуть только в контакте с дном мелководных водоёмов, где они оставались совершенно неэффективными необозримое время. В водной среде конечности намного менее функциональны, чем плавники и туловищная мускулатура. Очевидно, что требовались своеобразные условия, в которых функционирование явно более мощной и совершенной
туловищной мускулатуры стало бы менее эффективным, чем использование конечностей. В такой ситуации можно было бы ожидать, что конечности начнут быстро прогрессировать и увеличиваться, а туловищная и хвостовая мускулатура — редуцироваться. Этим условиям явно не удовлетворяет «прибрежно-мелководная» модель происхождения наземных позвоночных, где большинство двоякодышащих успешно передвигается при помощи змеевидных изгибов тела.
Палеонтологические свидетельства тоже предоставляют более чем скромную информацию на эту тему. У хорошо известных позднедевонских лабиринтодонтов (Ichthyostega) известны уже вполне полноценные конечности и предкрышечная кость, которая могла закрывать рудиментарные жабры. Это существо было явным хищником с развитым челюстным аппаратом и крупными зубами. Остаются открытыми вопросы о том, для чего ихтиостегам понадобились конечности и на кого они могли охотиться в конце девона. Если они охотились в воде на рыб, что наиболее вероятно, то конечности им только мешали. Вне водоёмов доступной для столь несовершенных тетрапод пищи было немного. Наземные беспозвоночные были не самыми медлительными и пассивными.
Биологические причины возникновения конечностей и выход на сушу хищных ихтиостег непонятны, тем более что ясных палеонтологических свидетельств о существовании промежуточных филогенетических звеньев
между рипидистиями и амфибиями пока не обнаружено. Отдельное внимание следует уделить преобразованиям органов чувств при выходе позвоночных на сушу.
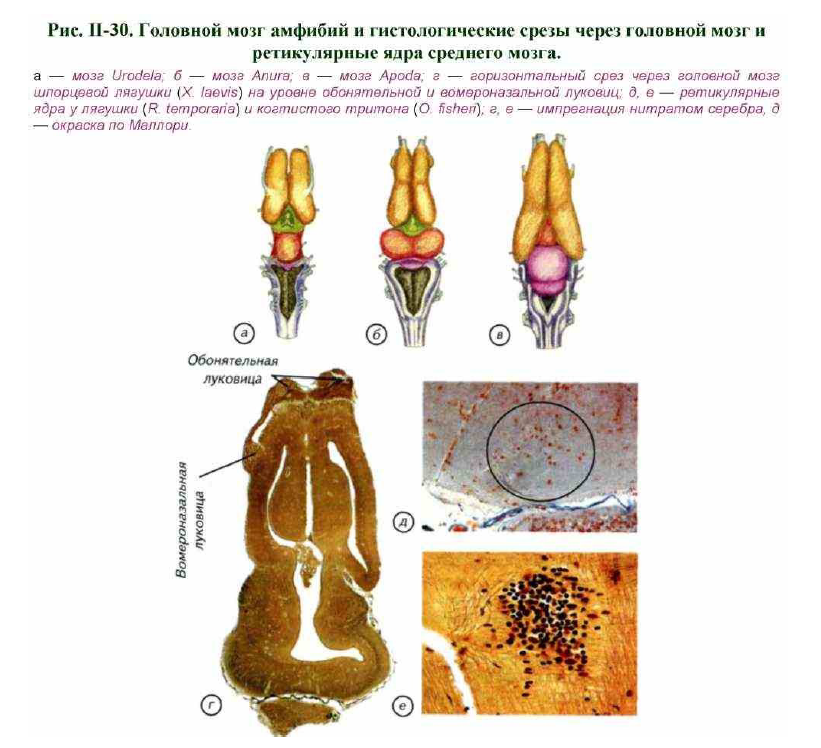
По мнению многих авторов, у примитивных тетрапод был хорошо развит орган обоняния (рис. II-30). Это вступает в противоречие с предположением о существовании переходных форм в мелких водоёмах, перенасыщенных разлагающимися остатками растений и животных. Довольно трудно представить себе биологический смысл развития обонятельной системы позвоночных в зловонных девонских
лужах. Развитие системы воздушного обоняния в таких условиях лишено всякого смысла. Возможны два решения этой проблемы: или обоняние не развивалось, или разлагающихся остатков было не настолько много, насколько нужно для стимуляции развития лёгочного дыхания. Развитие обонятельной системы у примитивных рипидистий и примитивных амфибий не вызывает сомнений, поэтому приходится отказываться от идеи возникновения лёгких в «зловонных, лишённых кислорода и пересыхающих водоёмах».
Ещё более парадоксальная ситуация возникает при анализе слуховой системы рипидистий. По общему мнению, лабиринт у этих животных развит намного лучше, чем у их потомков. Полукружных каналов три и они очень крупные. У примитивных амфибий это также хорошо развитый орган. Он включает в себя три органа чувств: слуховой аппарат (или его кожный аналог — боковая линия), рецептор линейного ускорения
(гравитационный рецептор) и рецептор углового ускорения (и торможения). Вполне справедлив вопрос о том, зачем такая совершенная система управления движением, если животное ползает в мелком водоёме или грязи миллионы лет. Если бы это было действительно так, то один из полукружных каналов редуцировался, как и у круглоротых. Поскольку этого не произошло, можно с уверенностью сказать, что обладатели такого развитого рецептора ускорения существовали в трёхмерной среде, а не барахтались в пересыхающих лужах. С этим выводом согласуются особенности строения специфической сенсорной системы, характерной для всех саркоптеригий. Рецепторы этой системы были расположены в
поверхностных слоях покровного скелета.
Под эмалеподобной поверхностью у древних рипидистий и двояко-дышащих располагается своеобразный дентин с системой поровых каналов и тонкими сенсорными порами. Найденные у рипидистий образования сходны по строению с ампулами Лоренцини и электрорецепторами современных хрящевых и
костистых рыб (Thomson, 1975; Vorobyeva, 1980). Эти органы необходимы для ориентации в темноте, где другие дистантные рецепторы малоэффективны. Акулы и костистые рыбы используют этот рецепторный орган на больших глубинах или в толще мутной воды. Учитывая совершенство морфологического строения ампулярного аппарата рипидистий, кажется вполне логичным, что порог их чувствительности был близок к порогу чувствительности отдельного рецептора в ампуле Лоренцини современных акул. Если даже его чувствительность была в несколько десятков раз ниже, это всё равно был очень совершенный орган, который позволял ориентироваться и искать добычу по искажениям электрического поля. Следовательно, саркоптеригий сформировали специализированную систему рецепции, для которой нужен довольно
большой объём воды.
В неглубоких и пересыхающих водоёмах такой дистантный анализатор практически бесполезен. По сути дела создаётся взаимоисключающая ситуация: наиболее совершенный рецептор возник и усовершенствовался в условиях, где его невозможно использовать. По-видимому, этот орган сформировался и использовался Crossopterygii в более подходящих условиях, чем пересыхающие водоёмы.
Довольно своеобразно выглядит организация зрительной системы саркоптеригий. У них обнаружены небольшие глазницы, которые свидетельствуют о незначительном развитии зрительной системы (рис II-31).
Это странная ситуация, поскольку в «мелководных и пересыхающих» водоёмах зрение должно играть доминирующую роль. Однако вопреки всем ожиданиям развитая зрительная система у рипидистий не встречается.
Среди разлагающихся органических остатков абсолютно «ненужная» обонятельная система быстро эволюционирует и дифференцируется в две системы с самостоятельной иннервацией и соответствующими центрами в головном мозге. Возникли две системы водно-воздушного обоняния. У амфибий наряду с основным органом обоняния появилась добавочная — вомероназальная (якобсонова) система дистантной хеморецепции.
Такимобразом, обоняние интенсивно эволюционировало, приспосабливалось к воздушной среде, а зрительная система изменялась незначительно.
Попробуем представить себе загадочного предка наземных позвоночных в широко признанной среде
обитания. Неглубокий, прогретый и периодически пересыхающий водоём так заполнен разлагающимися органическими остатками, что у рипидистий возникли лёгкие для воздушного дыхания. В этом водоёме при недостатке пищи находится слабовидящее животное, лишённое возможности использовать развитое обоняние, и практически глухое. Эту безрадостную картину дополняет ненужный в мелких водоёмах, но хорошо
развитый рецептор ускорения и такой же никчёмный, но совершенный дистантный электрорецептор.
Животное обладает всем, что ему совершено не нужно, и не имеет того, что ему просто необходимо. Все перечисленные противоречия приносятся в жертву одной идее — мелководной промежуточной среде обитания, которая считается «естественным» этапом между водной средой и сушей.
Таким образом, перечисленные особенности строения Crossopterygii ставят под сомнение гипотезу«прибрежно-мелководного» происхождения наземных позвоночных. Вероятность «счастливой случайности», позволившей преодолеть эти проблемы, крайне мала.

Следовательно, если мы хотим ясно представить события, приведшие к появлению тетрапод, необходимо ответить на ряд простых вопросов.
Почему предки позвоночных стали жить у побережья? Отчего возникла необходимость в воздушном дыхании? Почему слаборазвитые конечности оказались более эффективными, чем работа совершенной туловищной мускулатуры? Почему у примитивных четвероногих хорошо развиты обоняние, системы контроля движения и плохо развито зрение?
Ответы на эти вопросы легко получить, если перечислить биологические условия, для которых известная организация органов чувств и мускулатуры Crossopterygii и дошедших до нас останков древних амфибий (см. рис. II-30; II-31) была бы оптимальной. Водное, как и воздушное, обоняние имеет биологический смысл, если оно пригодно для поиска пищи, ориентации и размножения. Нам известно, что у амфибий
впервые сформировалась добавочная обонятельная система — якобсонов орган. В результате амфибиям стала доступна для химического анализа как воздушная, так и водная среда. Столь глубокие изменения органов обоняния, а не зрения, говорят о том, что зрительный анализатор был явно менее эффективным в тот период, хотя, казалось бы, в пересыхающих прибрежных водоёмах должно было бы преобладать зрение.
Следовательно, выход на сушу осуществлялся не через мелководные водоёмы, а в какой-то другой переходной среде.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы