22. Нервная система членистоногих
Организация нервной системы членистоногих и сходных с ними групп может существенно варьировать, но в пределах общего плана строения.
Рисунок нервной системы дневной бабочки (Lepidoptera) довольно точно отражает типичное расположение основных ганглиев в теле насекомых (см. рис. II-7, а). У насекомых самыми крупными являются головные ганглии. Они имеют довольно сложное строение.
Обычно в них выделяют несколько отделов (см. рис. II-7, е-и). Ростральнее всех расположены грибовидные тела, или corpora pedunculata.
Они являются высшим «интеллектуальным» центром беспозвоночных. По сути это функциональный аналог лобных долей большого мозга человека.
В грибовидных телах принимаются решения об использовании той или иной программы поведения. Более того, они организованы по принципам,общим с корой головного мозга млекопитающих. В коре и грибовидных телах нервные клетки расположены слоями или стратифицированы (см. рис. I-16).
Такой тип строения ассоциативных центров универсален для различных позвоночных и беспозвоночных животных. Он позволяет
оптимально организовать обработку распределённых потоков разнородных сигналов от специализированных сенсорных центров. Когда какой-либо отдел мозга позвоночных или ганглиев беспозвоночных специализируется на сложных анализаторных/ассоциативных функциях, он всегда формирует стратифицированные структуры (Беклемишев, 1964)
Грибовидные тела не имеют прямых связей с анализаторами. Только сигналы, предварительно обработанные в зрительных, вкусовых, обонятельных или соматических центрах, попадают в грибовидные тела. В них собирается и сравнивается уже интегрированная информация от внутренних и внешних рецепторов. Сравнение разнородных воздействий позволяет выбрать приоритетное и запустить максимально адекватную
программу поведения. Программ довольно много и они могут модифицироваться в зависимости от возникающих обстоятельств. Однако принципиальным изменениям они, как правило, не подвергаются.
Адекватность выбора программы поведения грибовидными телами — залог успеха или выживания конкретной особи. Для оптимального выбора принципиально важна предельная точность оценки внешних и внутренних сигналов, поэтому органы чувств многих беспозвоночных достигли феноменальной чувствительности, несопоставимой с возможностями позвоночных животных. В основе дистантной чувствительности
беспозвоночных лежат зрение, обоняние, механорецепция и различные формы слуховой чувствительности. В соответствии с развитием рецепторных систем различается и морфология головных ганглиев. К протоцеребруму относятся и грибовидные (стебельчатые) тела.
Протоцеребрум представляет собой зрительный центр, расположенный рострально (см. рис. II-7, ж; рис. II-8, г).
Дейтоцеребрум представляет собой хеморецепторный центр, в котором есть представительство обонятельных антенн (см. рис. II-7, з). Это представительство может различаться в десятки раз по объёму нервных
клеток.
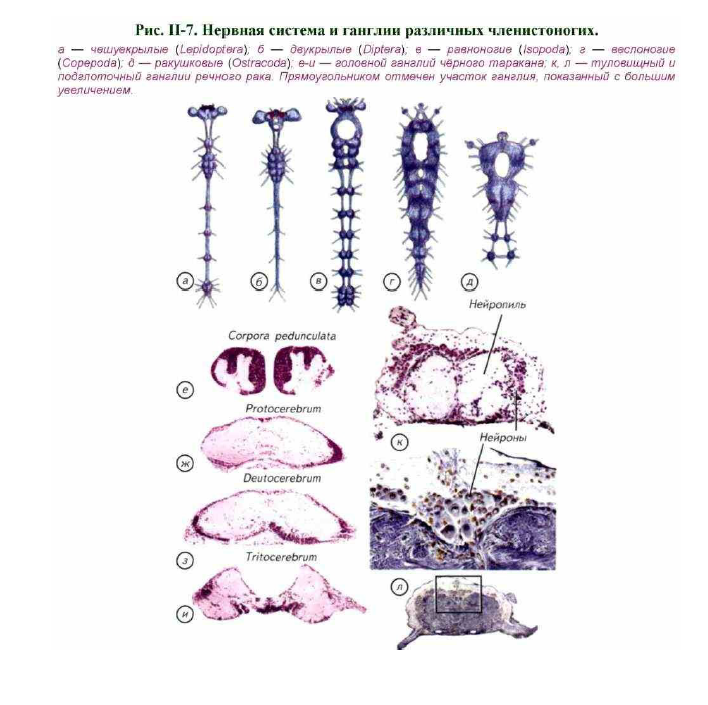
Однако если антенны у многих ночных бабочек имеют площадь поверхности примерно в 700 раз больше, чем у дневных, то различия по объёму дейтоцеребрума намного меньше. Тем не менее у хелицеровых вместе с редукцией антенн практически исчезает и дейтоцеребрум. Трито- церебрум выражен далеко не так хорошо, как два предыдущих отдела. Он связан преимущественно с работой нервов, иннервирующих верхнюю губу (см. рис. II-7, и). Поскольку ротовой аппарат членистоногих крайне изменчив, тритоцеребрум часто морфологически не выражен.
Подглоточный ганглий у всех членистоногих хорошо развит. Он иннервирует ротовые придатки и слюнные железы.
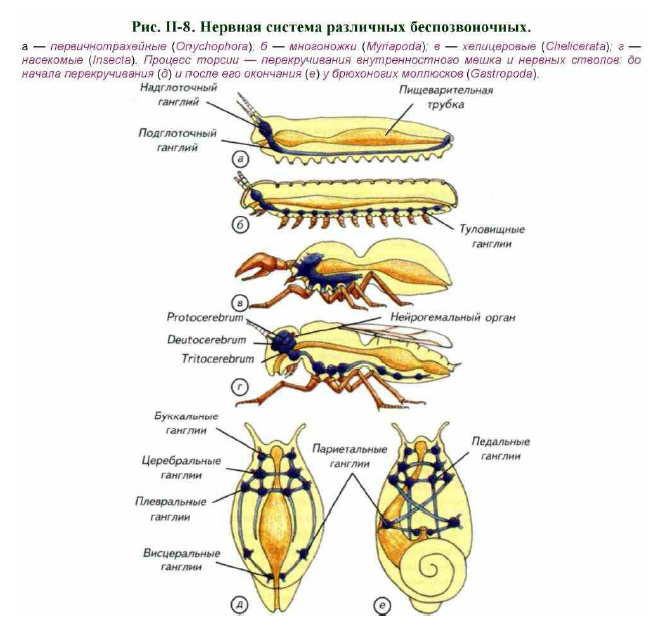
Этот общий план строения членистоногих был бы неполным без упоминания об нейроэндокринной системе. Основным центром секреции нейрогормонов у насекомых является нейрогемальный орган (см. рис. II-8, г). Это традиционное название в современной литературе несколько трансформировано. Под нейрогемальным органом понимают не всю гормональную часть головных ганглиев, а только его запасающий участок.
Гормональный комплекс расположен позади головных ганглиев и состоит из нескольких отделов. Большую часть этого комплекса составляет кардиальное тело, которое включает в себя железистую (секреторную) и запасающую части. В первой происходит синтез гормонов, а во второй — их накопление и по мере необходимости выделение. Кроме кардиального тела, к нейроэндокринным органам относят и небольшой парный ганглий,
называемый прилежащим телом. Он связан нервными волокнами как с кардиальным запасающим телом, так и с протоцеребрумом. Из ганглиев нейроэндокринного комплекса гормоны выделяются непосредственно в кровь или действуют опосредованно, влияя на эндокринные органы.
Примером может служить проторакальная железа, активность которой регулируется проторакотропным гормоном, выделяемым кардиальным телом. Эта железа выделяет экдизон, который регулирует начало линьки.
Примером прямого действия нервной системы на периферические органы является синтез диуретического гормона. Он синтезируется нейросекреторными клетками головных ганглиев и накапливается в запасающей части эндокринного отдела. Выделяясь из кардиального тела, он стимулирует выделение мочевины мальпигиевыми сосудами.
Нейрогормонов у членистоногих определено достаточно много, что вполне соответствует нашим представлениям о детерминационном принципе организации поведения животных этой группы. Её суть сводится к запуску определённой программы поведения в зависимости от состояния
организма или окружающей среды. При такой организации выбора стратегической направленности поведения гормональная регуляция является простым и эффективным способом управления всем организмом животного.
Если летающее насекомое отправляется в полёт, то вырабатывается адипокинетический гормон. Он мобилизует липиды организма как источник энергии для обеспечения полёта. Такой способ управления организмом построен на принципах обратной связи и по сути ничем не отличается от гормональной регуляции у млекопитающих.
Головные ганглии у членистоногих расположены над пищеварительной трубкой и соединены с подглоточным ганглием при помощи парных комиссур (см. рис. II-7; II-8). Подглоточный ганглий весьма разнообразен как по размерам, так и по клеточному составу. Даже у речного рака качественное разнообразие типов нейронов подглоточного ганглия сопоставимо с мозговыми структурами млекопитающих (см. рис. II-7, л).
От подглоточного ганглия начинаются отростки нейронов, иннервирующие ростральный участок пищеварительной трубки и ротовые придатки.
Контроль потребляемой пищи является одним из ключевых процессов, что отражается и на размерах этого нервного центра. Позади подглоточного нервного ганглия лежит нервная цепочка, состоящая из раздельных или интегрированных ганглиев (см. рис. II-7, а-д). Половая
система обычно иннервируется группой объединённых ганглиев в конце тела.
Различные формы организации ганглиев брюшной нервной цепочки обычно не сказываются на поведении членистоногих и сходных с ними групп. Крайне сложное инстинктивное поведение можно наблюдать как у общественных насекомых, так и у представителей ракообразных с сегментарно-лестничной организацией брюшной нервной цепочки. Таким примером могут быть представители равноногих ракообразных Isopoda)
(см. рис. II-7, в). Продольные нервные стволы и ганглии равноногих ракообразных полностью разделены, как и у плоских червей. Поперечные комиссуры образуют сходные «ступеньки» лестницы, но головные ганглии намного крупнее и дифференцированнее, чем у червей. В такой нервной системе достаточно полно сохраняется древнее сегментарное строение, но поведение представителей этой группы крайне разнообразно и не менее
сложно, чем у высших членистоногих. Сухопутные мокрицы, размножающиеся в норах, заботятся о своих потомках на протяжении 10 нед. Взрослые особи не выпускают молодь из норы и выкармливают её остатками растений. Процесс спаривания водяных осликов продолжается более 10 ч и осложнён переносом спермы при помощи шприцевидной конечности. Такая забота о потомстве, постоянные места для размножения и сложный половой процесс говорят о практическом равенстве возможностей членистоногих с различным строением нервной системы.
Концентрация головных и туловищных ганглиев членистоногих шла различными путями. Самым общим принципом стали укорочение комиссур и слияние ганглиев правой и левой стороны тела (см. рис. II-7, а- в). Этот процесс может сопровождаться рострокаудальным уменьшением количества нервных сегментов. В конечном счёте может происходить полная интеграция брюшной нервной цепочки до двух или одного анатомического центра: головных и грудных ганглиев. Среди насекомых своеобразным венцом совершенства организации такого типа являются двукрылые (Diptera). B этом отряде, как правило, брюшные нервные ганглии отсутствуют (см. рис. II-7, б). У десятиногих раков (Decapoda) хорошо выражены только головной и торакальный ганглии. При этом относительная длина продольных комиссур существенно меньше, чем у насекомых.
Однако латеромедиальное и каудоростральное слияние ганглиев может приводить к своеобразному строению нервной системы членистоногих и сходных с ними групп. Примером могут служить свободноживущие веслоногие ракообразные (Copepoda, Calanoida) (см. рис. II-7, г). Их головные и туловищные нервные ганглии сливаются в непрерывную структуру, которая мало напоминает нервную цепочку. Это связано с тем,
что у этих в основном мелких (1–6 мм) ракообразных голова сливается с грудью, а поведение очень незатейливо. Находясь в толще воды, они являются специализированными фильтраторами. В этой группе ракообразных сердце стало редкостью, а полостные жидкости передвигаются под механическим воздействием кишечника. Жабры обычно отсутствуют, а газообмен осуществляется через всю поверхность
тела. Об уровне специализации веслоногих ракообразных говорит то, что края их жующих ротовых пластинок усилены кремниевыми соединениями, не встречающимися у других высших членистоногих. Надо отметить, что нервная система свободноживущих веслоногих ракообразных намного сложнее, чем у представителей этой группы, питающихся придонными осадками или ведущих паразитический образ жизни. Для последних
характерно упрощение организации и уменьшение размеров всей нервной системы.
Самая высокая концентрация нервных ганглиев у ракушковых ракообразных (Ostracoda) (см. рис. II-7, д). Эти рачки обычно небольшого размера, у них утрачена сегментация. Тело рачка обычно заключено в двустворчатую хитинизированную и кальцифицированную раковину. Раковина часто может закрываться, а снаружи появляются только конечности. Биология этих ракообразных очень разнообразна. Среди них встречаются как фильтраторы, так и настоящие хищники. Они могут обитать и в морском грунте, и на суше, что говорит о всем организмом животного. Если летающее насекомое отправляется в полёт, то вырабатывается адипокинетический гормон. Он мобилизует липиды организма как источник энергии для обеспечения полёта. Такой способ управления организмом построен на принципах обратной связи и по сути ничем не отличается от гормональной регуляции у млекопитающих.
Головные ганглии у членистоногих расположены над пищеварительной трубкой и соединены с подглоточным ганглием при помощи парных комиссур (см. рис. II-7; II-8). Подглоточный ганглий весьма разнообразен как по размерам, так и по клеточному составу. Даже у речного рака качественное разнообразие типов нейронов подглоточного ганглия сопоставимо с мозговыми структурами млекопитающих (см. рис. II-7, л).
От подглоточного ганглия начинаются отростки нейронов, иннервирующие ростральный участок пищеварительной трубки и ротовые придатки. Контроль потребляемой пищи является одним из ключевых процессов, что отражается и на размерах этого нервного центра. Позади подглоточного нервного ганглия лежит нервная цепочка, состоящая из раздельных или интегрированных ганглиев (см. рис. II-7, а-д). Половая система обычно иннервируется группой объединённых ганглиев в конце тела.
Различные формы организации ганглиев брюшной нервной цепочки обычно не сказываются на поведении членистоногих и сходных с ними групп. Крайне сложное инстинктивное поведение можно наблюдать как у общественных насекомых, так и у представителей ракообразных с сегментарно-лестничной организацией брюшной нервной цепочки. Таким примером могут быть представители равноногих ракообразных Isopoda)
(см. рис. II-7, в). Продольные нервные стволы и ганглии равноногих ракообразных полностью разделены, как и у плоских червей. Поперечные комиссуры образуют сходные «ступеньки» лестницы, но головные ганглии намного крупнее и дифференцированнее, чем у червей. В такой нервной системе достаточно полно сохраняется древнее сегментарное строение, но поведение представителей этой группы крайне разнообразно и не менее
сложно, чем у высших членистоногих. Сухопутные мокрицы, размножающиеся в норах, заботятся о своих потомках на протяжении 10 нед. Взрослые особи не выпускают молодь из норы и выкармливают её остатками растений. Процесс спаривания водяных осликов продолжается более 10 ч и осложнён переносом спермы при помощи шприцевидной конечности. Такая забота о потомстве, постоянные места для размножения и сложный половой процесс говорят о практическом равенствевозможностей членистоногих с различным строением нервной системы.
Концентрация головных и туловищных ганглиев членистоногих шла различными путями. Самым общим принципом стали укорочение комиссур и слияние ганглиев правой и левой стороны тела (см. рис. II-7, а-в). Этот процесс может сопровождаться рострокаудальным уменьшением количества нервных сегментов. В конечном счёте может происходить полная интеграция брюшной нервной цепочки до двух или одного анатомического центра: головных и грудных ганглиев. Среди насекомых своеобразным венцом совершенства организации такого типа являются двукрылые (Diptera). B этом отряде, как правило, брюшные нервные ганглии отсутствуют (см. рис. II-7, б). У десятиногих раков (Decapoda) хорошо выражены только головной и торакальный ганглии. При этом относительная длина продольных комиссур существенно меньше, чем у насекомых.
Однако латеромедиальное и каудоростральное слияние ганглиев может приводить к своеобразному строению нервной системы членистоногих и сходных с ними групп. Примером могут служить свободноживущие веслоногие ракообразные (Copepoda, Calanoida) (см. рис. II—7, г). Их головные и туловищные нервные ганглии сливаются в непрерывную структуру, которая мало напоминает нервную цепочку. Это связано с тем, что у этих в основном мелких (1–6 мм) ракообразных голова сливается с грудью, а поведение очень незатейливо. Находясь в толще воды, они являются специализированными фильтраторами. В этой группе ракообразных сердце стало редкостью, а полостные жидкости передвигаются под механическим воздействием кишечника. Жабры обычно отсутствуют, а газообмен осуществляется через всю поверхность тела. Об уровне специализации веслоногих ракообразных говорит то, что края их жующих ротовых пластинок усилены кремниевыми соединениями, не встречающимися у других высших членистоногих. Надо отметить, что нервная система свободноживущих веслоногих ракообразных намного сложнее, чем у представителей этой группы, питающихся придонными осадками или ведущих паразитический образ жизни. Для последних характерно упрощение организации и уменьшение размеров всей нервной системы.
Самая высокая концентрация нервных ганглиев у ракушковых ракообразных (Ostracoda) (см. рис. II-7, д). Эти рачки обычно небольшого размера, у них утрачена сегментация. Тело рачка обычно заключено в двустворчатую хитинизированную и кальцифицированную раковину.
Раковина часто может закрываться, а снаружи появляются только конечности. Биология этих ракообразных очень разнообразна. Среди них встречаются как фильтраторы, так и настоящие хищники. Они могут обитать и в морском грунте, и на суше, что говорит о высокой адаптивности их нервной системы. Надо отметить, что у этих ракообразных встречается как партеногенетическое, так и половое размножение. Их половые органы крайне сложны в строении и использовании. Самцы должны ввести свой половой аппарат под панцирь самки и только потом выпустить сперматозоиды в защищённую полость.
После оплодотворения самка обычно откладывает яйца в какое-либо укрытие. Ракушковые ракообразные проявляют своеобразную заботу о потомстве, поскольку оплодотворённые яйца могут развиваться до вылупления под раковиной матери. Все половые процессы у ракушковых ракообразных невероятно затруднены тем, что у многих видов длина сперматозоидов в несколько раз больше длины тела. Так, у представителей
рода Pontocypris сперматозоид в 8 раз длиннее тела взрослой особи и достигает 6 мм. Ракушковые ракообразные являются своеобразными сексуальными рекордсменами, поскольку и относительные, и абсолютные размеры их сперматозоидов не имеют аналогов среди животных.
Таким образом, у членистоногих и сходных с ними групп мы видим значительное разнообразие анатомического строения брюшной нервной цепочки и головных ганглиев. Однако они вписываются в общий архетип и обеспечивают сложное инстинктивное поведение. Поведение свободноживущих форм как при лестничной нервной системе, так и при единственном цереброторакальном ганглии существенно не различается. В обоих случаях мы видим многоступенчатые инстинкты, заботу о потомстве, сложное пищевое и половое поведение. Структурная организация головных ганглиев и брюшной нервной цепочки даёт идеальную возможность для создания универсальной системы с запрограммированным, но адаптивным поведением.
Брюшной нервной цепочкой далеко не исчерпываются варианты строения нервной системы беспозвоночных. Существует множество групп этих животных, достойных рассмотрения как с позиций строения нервной системы, так и в связи со сложностью поведения. Весьма демонстративна организация моллюсков (Mollusca). У этих беспозвоночных встречаются самые контрастные формы строения нервной системы с невероятным
разнообразием поведения. У моллюсков известны примитивнейшие нервные системы, состоящие из нескольких асимметричных ганглиев или вообще без них. Такой тип нервной системы характерен для малоподвижных или прикреплённых животных, и среди моллюсков есть настоящие рекордсмены. Самое крупное неподвижное животное на этой планете — тридактна (T. gigas) достигает вместе с раковиной массы более 200 кг. Хорошо известны и свободноживущие брюхоногие (Gastropoda) и головоногие (Cephalopoda) моллюски со сложным инстинктивным поведением и способностью решать сложные экспериментальные задачи, не встречающиеся в естественных условиях обитания.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы