Глава III. Становление мозга амниот
34. Репродуктивные стратегии амниот
К амниотам относят рептилий, птиц и млекопитающих. Группа объединена по эмбриологическому принципу устройства оболочек зародышей. Они принципиально отличаются от неклеточных икряных мешков или стенок икринок амфибий (Ромер, Парсонс, 1992). У амфибий размножение обычно связано с водой, а внезародышевые оболочки отсутствуют. Икринки откладываются самками амфибий в воду, где они оплодотворяются сперматозоидами самцов. В икринках развиваются эмбрионы, которые трансформируются в личинки и переходят к самостоятельному питанию в воде (рис. III-1).
Затем следует метаморфоз с выходом на сушу или половое созревание для водных и неотенических форм амфибий. Оплодотворение созревших яйцеклеток в воде является древнейшим способом размножения, характерным для первичноводных позвоночных. У многих амфибий происходит внутреннее оплодотворение, хотя эмбрионы и личинки по- прежнему развиваются в воде (см. рис. III-1; рис. III-2).
По-видимому, у лабиринтовых предков амниот размножение отличалось от архаичных рыб (Dodd, Dodd, 1985). Первоначально они могли иметь наружное оплодотворение и откладывать икру в укромных местах своих охотничьих лабиринтов. Однако риск уничтожения потомства в таких условиях очень велик. Снизить гибель потомства можно было только одним способом — носить икру с собой. Появление такой заботы о потомстве довольно обычно у костистых рыб и неоднократно происходило в процессе эволюции (Jameson, 1988).
Забота о потомстве должна была неизбежно привести к появлению внутреннего оплодотворения с последующим вынашиванием развивающейся икры внутри тела (см. рис. III-1; III-2).
Переход к живорождению стал у лабиринтовых амфибий принципиальным шагом на пути адаптации к наземной среде. Размножение стало отчасти независимым от водной среды. У архаичных амфибий существовали все возможные варианты наружного и внутреннего оплодотворения, длительного или краткосрочного вынашивания развивающейся икры (см. рис. III-2, а-г). Наиболее успешными стали животные с длительным вынашиванием зародышей. Судя по всему, они могли появляться на свет уже полностью готовыми к самостоятельному существованию. При откладке икры в воду зародыш может иметь минимальный запас желтка (см. рис. III, а). Личинка рано выходит из икринки и начинает питаться самостоятельно. При удлинении срока развития потребность в желтке или другом источнике питательных веществ резко возрастает.
Эта проблема решается несколькими способами. Можно использовать индивидуальный запас желтка или богатые секретом яйцеводы материнского организма. Зародыши некоторых живородящих пластиножаберных питаются стенкой яйцеводов вплоть до выхода из материнского организма. Однако чаще всего просто увеличивается индивидуальный запас желтка, в результате зародыш может долго находиться в организме матери. Он появляется на свет уже сформированным организмом, готовым к самостоятельному питанию.
Такие варианты заботы о потомстве часто встречаются во всех группах первичноводных позвоночных. Ни к каким особым эволюционным достижениям такое внутреннее развитие, казалось бы, привести не должно.
Тем не менее одна особенность биологии отличает древних амфибий от первичноводных позвоночных. Амфибийный образ жизни предусматривает пребывание на суше, где механические деформации тела под действием гравитации заметнее, чем в водной среде. Механические воздействия на репродуктивные органы, ещё не защищённые специализированными элементами скелета, неизбежно должны были привести к развитию компенсаторных демпферов — дополнительных оболочек (см. рис. III-1, б). Если бы зародыш находился вне организма матери, то никаких специальных внезародышевых оболочек не понадобилось, но на суше деформационные воздействия на органогенез внутри яйцеводов весьма велики.
Это связано с тем, что на ранних стадиях развития позиционная информация в морфогенетически активных зачатках эмбриона кодируется при помощи механогенетических процессов. Суть этого явления состоит в изменении проницаемости механозависимых ионных каналов при формообразовании. Они обеспечивают каждой клетке индивидуализированный позиционный сигнал, который биомеханически интегрирован со всем зачатком или растущим органом. Благодаря этим взаимодействиям клетка периодически получает управляющий позиционный сигнал и запускает региональные гистогенетические процессы (Савельев, 1993).
Надо отметить, что этот механизм регуляции раннего эмбрионального формообразования характерен для всех
позвоночных и предопределяет необходимость биомеханической защищённости развития. Понятно, что столь ненадёжный механизм весьма уязвим. Даже при незначительных статичных деформациях зародыша легко возникают множественные аномалии развития и эмбрион погибает.
Любой переход к внутреннему развитию икринок должен предусматривать снижение механической нагрузки на зародыш. У первичноводных позвоночных сама среда создаёт биомеханически благоприятные условия для внутреннего развития икринки. Зародыш, как и материнский организм, находится в механически компенсированной водной среде. При переходе на сушу это условие перестаёт соблюдаться.
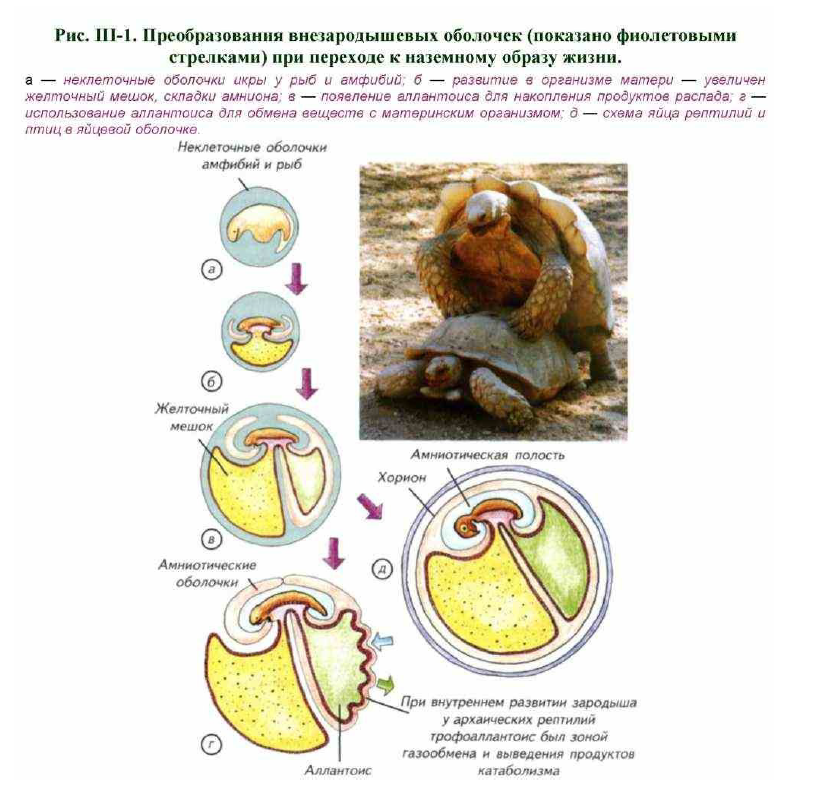
При переходе на сушу это условие перестаёт соблюдаться. Обойтись простои неклеточной оболочкой уже практически невозможно. Неизбежно начинают формироваться внезародышевые оболочки (см. рис. III-1, а-в).
Первоначально они возникают как демпферные структуры, окружающие зародыш. Самым простым способом механической защиты зародыша в яйцеводе становятся увеличение количества желтка и создание многослойной оболочки вокруг зародыша (см. рис. III-1, в, г). Судя по всему, амниотические оболочки и желточный мешок формировались в эволюции параллельно. Амниотические оболочки происходят из эктодермы, а желточный мешок — из энтодермы. Через амниотические оболочки обеспечивается обмен амниотической жидкости, омывающей зародыш. Желточный мешок служит для хранения питательных веществ зародыша, является источником первичных половых клеток и содержит клетки-предшественники форменных элементов крови.
Чем больше становился зародыш при внутреннем развитии архаичных рептилий, тем актуальнее становилась утилизация продуктов метаболизма эмбриона. Внутри тела матери пространство вокруг зародыша замкнуто, что требует некоего хранилища отходов или способа их выведения. На первом этапе эволюции внезародышевых оболочек эта проблема была решена просто: сформировался дополнительный мешок — аллантоис Это энтодермальное выпячивание вентральной поверхности эмбриональной задней кишки.
При развитии зародыша в аллантоисе накапливаются мочевина, мочевая кислота и азотистые продукты обмена. Любой из этих компонентов может летально изменить осмотический баланс вокруг зародыша, поэтому они изолированы друг от друга. Однако в аллантоисе не только накапливаются продукты жизнедеятельности зародыша. Через него у рептилий и птиц происходит газообмен при помощи специализированных кровеносных сосудов. У современных амниот все перечисленные оболочки окружены хорионом, или серозой, которая в зоне слияния с аллантоисом носит название хориоаллантоиса. Она также участвует в газообмене у современных рептилий и птиц.
Современные рептилии и птицы откладывают яйца в оболочках или обладают живорождением (см. рис. III-2, г, д). Дальнейшее развитие этих оболочек у млекопитающих привело к появлению хориоаллантоисной плаценты. Через неё происходит не только дыхание зародыша, но и обмен веществ, синтез гормонов и многие другие процессы (см. рис. III-2, е).
Следует отметить, что у многих современных рептилий выявлена примитивная плацентация (Stewart, Blackburn, 1988). Она не столь совершенна, как у млекопитающих, но имеет ряд свойств, позволяющих использовать её для питания развивающихся зародышей рептилий.
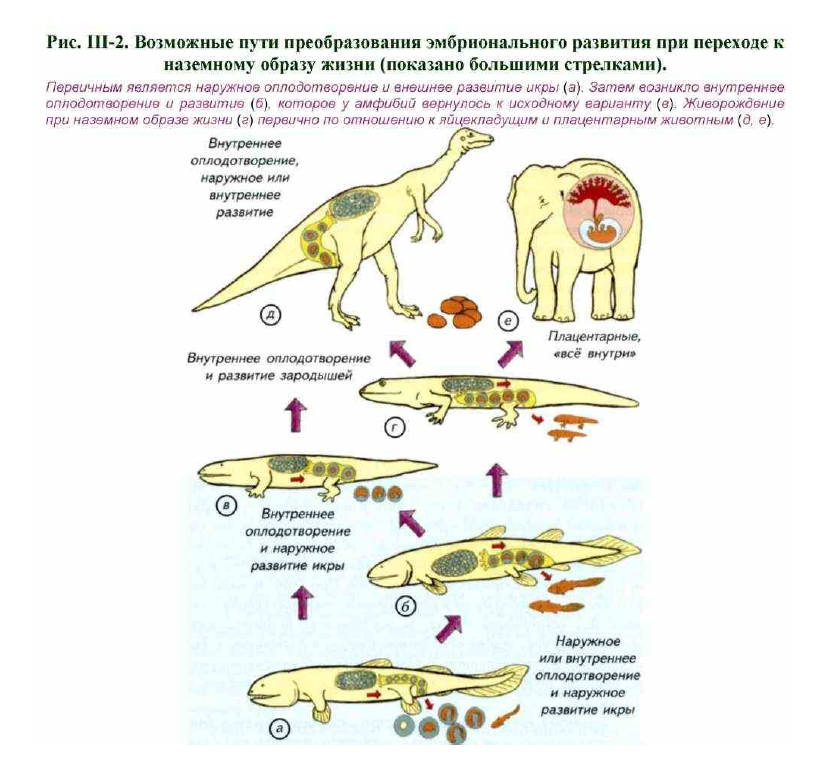
Происхождение амниотических оболочек у зародышей архаичных рептилий может быть обусловлено внутренним оплодотворением и развитием. На это указывает отсутствие в палеонтологической летописи убедительных доказательств способности примитивных рептилий откладывать яйца на протяжении всего палеозоя, что позволяет допустить несколько иной вариант развития событий.
На первом этапе возник вариант внутреннего оплодотворения с непродолжительным вынашиванием икры в родительском организме. В этот период дополнительные внезародышевые оболочки были развиты слабо (см. рис. III-1, a; III-2, б, в). В среде лабиринтов удлинение времени внутреннего развития привело к различным формам живорождения.
Следствием этого стало появление внезародышевых оболочек, которые создавали вокруг зародыша стабильную среду, обеспечивали запасом пищи и отводили продукты катаболизма. Пока зародыш был незначительных размеров, такое решение было очень успешным.
Архаичные рептилии вынашивали своё потомство внутри себя до рождения сформированных особей. При этом не надо было решать сложную проблему яйцевых оболочек, терморегуляции и газообмена. Все компоненты успеха, за исключением внезародышевых оболочек, были унаследованы от первичноводных позвоночных, которые обладают живорождением. С такой формой размножения уже не было никакой необходимости в водоёмах или особых местах для откладки яиц.
По-видимому, древние рептилии быстро расселились на суше благодаря живорождению, а не развитию способности откладки яиц с прочными оболочками (см. рис. III-2, г). Косвенным свидетельством в пользу этого вывода являются ихтиозавры, которые обитали в водной среде около 220 млн лет назад. Эти вторичноводные рептилии сохранили воздушное дыхание и были живородящими. Вынашивание в материнском организме крупных эмбрионов могло стать причиной появления специализированных участков оболочек, которые обеспечивали газообмен и отвод продуктов метаболизма, что известно для многих современных видов рептилий (Stewart, Blackburn, 1988).
Специализированный участок аллантоиса обеспечивал газообмен, а скопившиеся продукты жизнедеятельности зародыша могли выходить в материнский организм по законам осмоса. Увеличение складчатости внезародышевых оболочек привело к повышению площади обмена, а затем и к появлению прообраза плаценты — трофобластического аллантоиса, или трофоаллантоиса.
Трофоаллантоис был аналогом плаценты только в отношении газообмена. Кислород поступал к зародышу, а углекислый газ выводился в материнский организм. Продукты метаболизма зародыш передавал матери через трофоаллантоис, а питательные вещества получал из запасов желтка.
Следовательно, значительная часть рептилий могла обладать смешанным желточно-трофоаллантоисным обменом зародышей при сохранении внутреннего развития. Такой вариант давал заметные биологические преимущества архаичным рептилиям. Появлялась неограниченная возможность перемещений без привязки к местам размножения.
Сформировалась дополнительная защита зародыша при его внутреннем развитии. Организм матери гарантировал активную терморегуляцию, мать могла вынашивать крупных потомков, которые становились конкурентными сразу после рождения. Внутреннее оплодотворение привело к перестройке половой системы и полового поведения, что повлекло за собой развитие особых центров мозга.
Появление яйцекладущих рептилий явно было следующим этапом эволюции амниот (см. рис. III-2, д). В независимом от материнского организма яйце нет необходимости создавать многослойные демпферные оболочки и механически защищать зародыш. Если бы яйцекладущие рептилии появились, минуя этап внутриутробного развития, то следовало бы ожидать намного более упрощённого варианта строения внезародышевых оболочек. Сложные оболочки у всех амниот косвенно подтверждают существование продолжительного этапа внутреннего развития зародышей, который возник задолго до появления яиц с твёрдой оболочкой или плаценты млекопитающих (Van Tienhoven, 1983).
Плацента млекопитающих представляет собой вариант трофоаллантоиса, где функции газообмена и экскреции продуктов метаболизма дополнены поступлением питательных веществ через
разветвлённую сосудистую сеть.
Собственно говоря, плацента является трофоаллантоисом с расширенными функциями (Wimsatt, 1974). У большинства млекопитающих сохранён желточный мешок, хотя и в рудиментарном виде. Появление механизмов питания зародыша за счёт организма матери принципиально отличается от трофоаллантоисного типа развития. У млекопитающих впервые с помощью плаценты были сняты ограничения, связанные с размерами зародыша. Он мог быть любого размера, который смогла бы обеспечить ему мать. Источником
питательных веществ стал не ограниченный желточный мешок, а активный организм матери.
Таким образом, причиной появления внезародышевых оболочек было внутреннее развитие эмбрионов у амфибий и архаичных рептилий, вышедших на сушу. Они являются биомеханической защитой ранних этапов развития, хранилищем питательных веществ и местом накопленияпродуктов метаболизма. У архаичных рептилии происходили внутреннее оплодотворение и развитие эмбрионов. Это стало причиной формирования трофоаллантоисного обмена и расселения рептилий. Яйцекладущие рептилии возникали неоднократно, но уже после трофоаллантоисного этапа эволюции. Вторично возникшая возможность не вынашивать
зародыши, а откладывать яйца связана у рептилий с развитием специальных прочных наружных оболочек. Зародыши стали развиваться в замкнутой среде яйца, как в индивидуальном изолированном водоёме.
Биологическая вместимость такого персонального водоёма ограничена, а зависимость от внешней среды довольно велика. По-видимому, преобладание яйцекладущих рептилий было одной из причин их вымирания. Однако из трофоаллантоисной системы питания зародышей рептилий возникла плацента млекопитающих. С её помощью были сняты заложенные в объёме желтка ограничения на размеры зародыша и время вынашивания плода.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы