14. Эффекторные системы
В основе эффекторных систем лежит всё та же способность клеток воспринимать сигналы химической, механической и электромагнитной природы. Однако источником этих сигналов становится не внешняя среда, а сама нервная система. Внутренние органы и скелетная мускулатура получают управляющие сигналы от нервной системы или её посредников.
Изменение состояния этих эффекторных систем приводит к физической модификации наблюдаемого поведения. Надо отметить, что у животных и человека нет переизбытка вариантов воздействия на внешнюю среду.
Эффекторные системы включают в себя несколько компонентов. Основным реальным способом воздействия на внешний мир является механическая активность животного. Этот непосредственный эффекторный механизм построен на различных вариантах деформации тела. Тело можно перемещать в пространстве по гидродинамическим законам, как это делают кальмары, выбрасывая порции воды из-под мантии. Неплохой вариант аппарата движения представляют гидростатические амбулакральные ножки, которые используют морские звёзды. Однако основной способ двигательной активности у животных связан с сокращением мускулатуры.
Скелетная мускулатура состоит из нескольких типов поперечнополосатых мышц, которые различаются по величине, продолжительности усилий и типу метаболизма. Поперечная исчерченность этих мышц обусловлена упорядоченным расположением слоёв актиновых и миозиновых филаментов. Филаменты способны к движению относительно друг друга с затратой энергии, что и является основой механической активности мышц. Сходное строение имеет и сердечная мышца, но между её клетками расположены электрические соединения. Они позволяют распространяться электрическому току от клетки к клетке.
В скелетной мускулатуре обычно выделяют тонические, медленные и быстрые фазические волокна. Тонические мышечные волокна не могут совершать одиночных сокращений и сокращаются медленно. Их сокращение полностью контролируется нервными импульсами мотонейронов. Эти мышцы распространены у амфибий, рептилий и птиц.
У млекопитающих они встречаются только в наружных мышцах глазного яблока. Фазические мышечные волокна могут производить одиночные сокращения. Сигнал, пришедший к ним по нервному волокну, вызывает генерацию собственного мышечного электрического разряда — потенциала действия. Этот разряд распространяется по мышце и синхронизирует сокращение отдельных мышечных волокон. Медленные мышечные фазические волокна иннервируются одним нервным волокном и сокращаются относительно медленно. Они эффективно участвуют в поддержании позы животных, медленно «устают» и характеризуются окислительным типом обмена. Это означает, что в таких мышцах много белка миоглобина, связывающего кислород и придающего тёмно-красный цвет мясу рыб и птиц.
Похожий тип обмена выявлен и у быстрых фазических волокон. Это также окислительные волокна, но способные к быстрым одиночным сокращениям. Энергичные и повторяющиеся движения крыльев птиц служат хорошей иллюстрацией такой локомоции. Совершенно иной тип обмена свойствен быстрым фазическим волокнам. Они способны развивать огромные усилия, но крайне непродолжительно. Они быстро «устают» из-
за того, что энергия, необходимая для сокращения, продуцируется в результате гликолиза. Возникает торможение обмена кислорода, которое требует времени для восстановления исходной ситуации. Такие мышцы используются животными для экстремальных в силовом отношении ситуации. Гликолитические волокна широко распространены у амфибий, рептилий и птиц. Преимущества такой мускулатуры для холоднокровных
животных очевидны. Она позволяет развить большие усилия при низком уровне обмена. При взлёте птицы используют мощные грудные мышцы.
Они белого цвета и являются самым наглядным примеромгликолитической мускулатуры. Таким образом, эффекторная система скелетной мускулатуры весьма разнообразна и имеет несколько типов волокон. Однако смысл работы любых механических систем сводится к перемещениям тела животного или человека.
Для многих первичноводных позвоночных характерен ещё один эффекторный механизм, связанный с изменением окраски тела (рис. I-20).
Ещё в 1876 г. Пуше (G. Poushet) заметил, что при перерезке зрительного нерва меланофоры расширяются на поверхности тела карпозубых рыб и замирают в таком состоянии. Иначе говоря, он установил факт центральной нервной регуляции меланофоров костистых рыб и её зависимость от зрительной системы. Центр управления меланофорами расположен в вентральной части среднего мозга. От него начинается
эффекторный спинномозговой пучок волокон, который оканчивается на уровне XV–XVI позвонков. Через два или три вентральных корешка спинномозговые симпатические нервные волокна выходят в симпатический ствол и распространяются по всему телу животного.
Отростки двигательных симпатических нейронов достигают конкретных меланоцитов, что позволяет вызывать быстрые реакции по изменению окраски животного. На поверхности головы меланофорами управляютсимпатические волокна, которые выходят из головного мозга в составе тройничного (V) черепно-мозгового нерва, поэтому первичные реакции меланофоров костистых рыб на голове и теле существенно различаются как по скорости изменения, так и по цветовой гамме. Граница между двумя бассейнами иннервации проходит по жаберной крышке.
Кроме скелетных мышц, эффекторными системами животного являются гладкая мускулатура, эндокринные и экзокринные железы. Работа этих органов не так очевидна, как скелетной мускулатуры, но они оказывают огромное влияние на поведение животных. Гладко-мышечные клетки не имеют поперечной исчерченности, как сердечная мышца или скелетная мускулатура. Внутри цитоплазмы миофиламенты распределены относительно случайно. Гладкомышечные клетки расположены у позвоночных в стенках артерий, артериол, пищеварительной системы, мочеточников, мочевого пузыря и многих других систем. Этот эффекторный путь регулирует давление крови, прохождение пищи и активность половых органов. Механизм регуляции сокращения гладких мышц зависит от их близости к нервным окончаниям. Чем они ближе расположены, тем активнее реакция сокращения. Выход медиатора из нервных окончаний может модулировать активность гладко-мышечных клеток. Этот тип клеток чувствителен как к нервной, так и к гормональной регуляции. Длительные сокращения могут вызываться как нервной активацией, так и гормональными причинами.
Ещё более инерционным эффекторным механизмом управления поведением животного являются эндокринные железы (см. рис. I-20), это мощная система регуляции физиологической активности животного. Нервная система может на них влиять непосредственно или через нейроэндокринные клетки специализированного отдела — промежуточного мозга. В нём находятся гипоталамус, эпиталамус, эпифиз и гипофиз. Нейрогормоны этой части мозга контролируют практически все железы внутренней секреции. Исключение составляют паращитовидные железы и кальцитониноциты щитовидной железы, которые контролируют кальциевый обмен позвоночных. Их активность регулируется по принципу обратной связи в зависимости от концентрации кальция в плазме крови.
Рис. I-20. Эффекторные системы организма позвоночных.
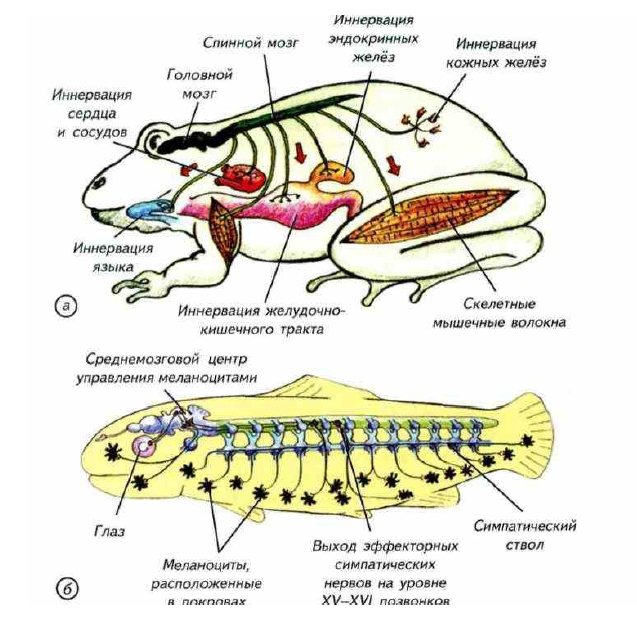
а — эффекторные нервные волокна управляют работой клеток гладкой и поперечнополосатой мускулатуры, наружных секреторных систем и желёз внутренней секреции; б — в покровах многих первичноводных позвоночных расположены пигментные клетки, которые меняют окраску животного в зависимости от окружающей среды. Эти клетки имеют центральную нервную регуляцию, а управляющий центр связан со зрительной системой животного.
У животных эндокринные железы вырабатывают различные химические вещества, которые переносятся от одних органов к другим.
Среди этих веществ есть соединения, влияющие практически на все системы организма и дополняющие действие нервной системы. Во многих случаях нервная система, стимулировав гормональную регуляцию поведения, сама оказывается под влиянием гормональной активности. По сути поведение животного контролируется двумя главными интегративными системами организма — нервной и эндокринной, которые тесно связаны между собой. Основной функциональной единицей эндокринной системы является гормон.
Гормонами называют особые химические вещества, вырабатываемые в одних и действующие в других частях организма. В большинстве случаев гормоны образуются в хорошо обособленных железах или отдельных органах. Воздействие гормона зависит от его химических свойств иприроды реагирующих тканей. Такие ткани названы тканями-мишенями.
Гормоны вырабатываются в щитовидной железе, мозговом веществе и коре надпочечников, яичниках, семенниках, аденогипофизе, нейрогипофизе, урогипофизе и в нервной системе. Выделение гормонов нервными клетками называется нейросекрецией.
Физиологическое подразделение гормонов на кинетические, метаболические, морфогенетические и поведенческие построено на их эффектах. Кинетические гормоны запускают мышечное сокращение, секреторную активность эндокринных и экзокринных желёз и изменение пигментации животных. Метаболические гормоны регулируют водно- солевой баланс, обмен углеводов и белков. Морфогенетические гормоны действуют в эмбриональный период, во время роста и полового созревания животных. Они регулируют рост тела, линьку, метаморфоз, созревание гонад, освобождение гамет и половую дифференцировку. Поведенческие гормоны являются достаточно условной группой, поскольку все гормоны
так или иначе влияют на активность животных. Тем не менее традиционно называют «поведенческими» гормоны, регулирующие сезонную половую активность, и гормоны, оказывающие трофическое действие на нервнуюсистему.
Гормональная регуляция поведения играет очень большую роль в повседневной и сезонной активности животных. Давно обнаружена прямая зависимость между отдельными гормонами и конкретными формами поведения. Гормональное воздействие на особь продолжительное и охватывает как нервную систему, так и организм в целом. Таким образом, гормоны влияют на поведение в большей степени, чем любая другая
система организма, за исключением мозга. Наибольшему гормональному контролю подвержены половое созревание, половой диморфизм, размножение, ухаживание, копуляция и инверсия пола. Тем не менее это только одна из эффекторных систем, регулирующих физиологическую активность и поведение животных.
Ещё одной эффекторной системой можно назвать экзокринные железы. Они участвуют в пищеварении, защите кожи, маркировании территории и половых контактах между особями. Вырабатываемые ими соединения выделяются во внешнюю среду, а не внутрь организма. При помощи кожных желёз многие рыбы и амфибии защищают поверхность тела от различных неблагоприятных воздействий, а экзокринная часть
поджелудочной железы вырабатывает пищеварительные ферменты. Работа глазничных и анальных желез млекопитающих служит одним из важнейших способов внутривидовой и межвидовой коммуникации.
Следовательно, нервная система реализует свою активность через мышечные клетки и железы. Они обеспечивают механизм движения самого животного, перемещение пищи и крови внутри его организма.
Прямо или косвенно эти эффекторные системы осуществляют управление и контроль за работой почти всех специализированных органов и функциональных систем организма. Результаты этой работы постоянно контролируются внутренними рецепторами и органами чувств.
Эффекторные и рецепторные структуры объединяет нервная система. Она прямо или косвенно влияет на весь организм, определяет его способность к быстрым адаптациям и контролирует взаимодействия между различными органами.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы