7. Гематоэнцефалический барьер.
Нервная ткань — это объединение специализированных клеток, которые воспринимают, обрабатывают, хранят и используют информацию о внешней среде и внутреннем состоянии организма. Этим функциям подчинено строение нервных клеток — нейронов. Нервные клетки имеют особенности, которые отличают их от других клеток организма (рис. I-9). Нейроны неодинаковы. Они различаются по размеру, форме ветвления дендритов и аксонов, выделению различных химических веществ и физиологической активности.
Рис. I-9. Строение нейронов и глиальных клеток.
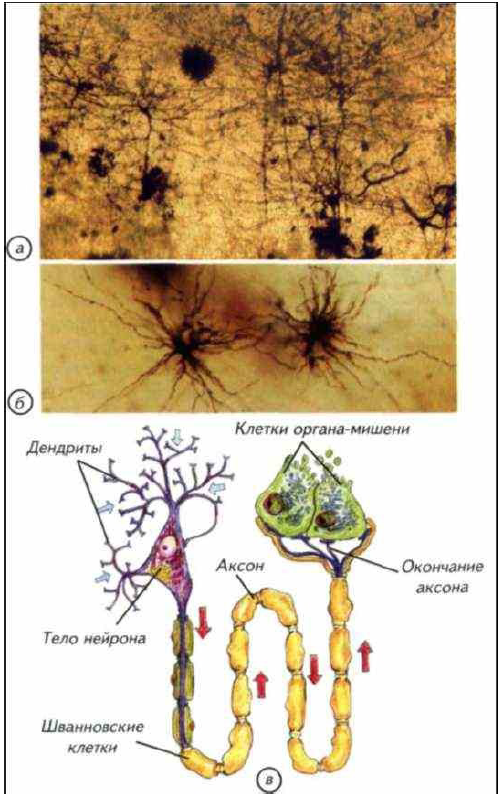
а — импрегнированные нейроны коры головного мозга человека. При такой окраске виден примерно 1 нейрон из 1000, что позволяет рассмотреть его отростки; б — глиальные клетки из мозга человека; в — строение нейрона и его аксона, закрытого глиальными клетками.
Нейроны — характерные структурные элементы нервной системы объединены в сети и в специализированные структуры ганглии или мозг, а их отростки образуют периферические нервы.
В нервных клетках — нейронах — обычно можно выделить клеточное тело, дендриты и аксон (см. рис. I-9). Тело содержит ядро и биохимический аппарат синтеза молекул, необходимых для жизнедеятельности клетки. Обычно тело нейрона имеет округлую, веретеновидную или пирамидальную форму. Дендриты представляют собой тонкие отростки, которые многократно ветвятся в непосредственной близости от тела клетки. Вокруг него образуется ветвистое дерево. Дендриты формируют ту основную физическую поверхность, на которую поступают идущие к данному нейрону сигналы. Аксоны распространяются далеко от тела клетки. Их длина варьирует от 1 мм до 1,5 м, что позволяет аксонам выполнять функции линий связи между телом клетки и далеко расположенным органом-мишенью или отделом мозга. По аксону проходят сигналы, генерируемые в теле данной клетки. Аксон отличается от дендритов как по строению, так и по свойствам наружной мембраны. Большинство аксонов длиннее и тоньше дендритов и имеют отличный от них характер ветвления. Отростки дендритов в основном группируются вокруг клеточного тела, тогда как отростки аксонов располагаются на конце волокна, в том месте, где аксон взаимодействует с другими нейронами или органами-мишенями.
Кроме нейронов, в нервной системе есть и другие специализированные клетки, которые не выполняют перечисленных нервных функций. Это клетки глии. Глиальные клетки не могут генерировать или обрабатывать информационные сигналы. В их задачу входят снабжение нейронов соединениями, необходимыми для нормального метаболизма, отведение продуктов катаболизма и обеспечение барьерных функций между мозгом и кровеносной системой. Кроме этого, глиальные клетки выполняют функции макрофагов, лимфоцитов и других клеток кровеносной и лимфоидной систем. Нейроглия выполняет механическую функцию и изолирует электрохимически активные волокна отдельных нервных волокон внутри мозга. Оболочки вокруг отростков нейронов состоят из клеток нейроглии, что позволяет стабилизировать ионную среду и увеличивать скорость проведения нервного сигнала (рис. I-10; I-11).
В головном мозге изолирующие функции выполняет олигодендроглия. Она происходит из нейроэктодермы, но отличается от нейронов тем, что не генерирует никаких сигналов, а специализируется на изолирующих функциях. Каждая клетка олигодендроглии охватывает сразу несколько отростков нейронов (см. рис. I-9). Олигодендроглия окружает отростки нейронов, тогда как другие глиальные клетки изолируют тела нейронов.
Рис. I-10. Основные компоненты гематоэнцефалического барьера головного мозга и периферической нервной системы.
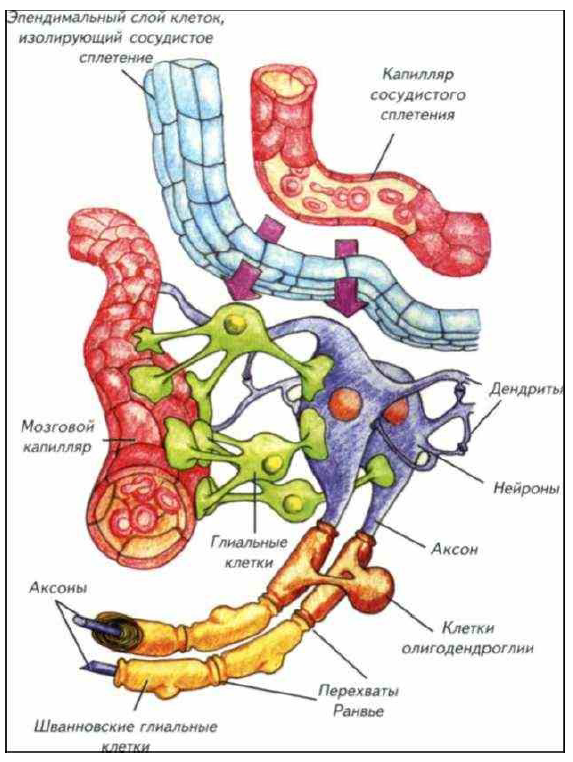
Головной мозг изолирован от кровеносной системы трофическими глиальными клетками (зелёные), олигодендроглией и шванновскими клетками. Спинномозговая жидкость фильтруется через эпендимные клетки нейрального происхождения.
Глиальные клетки выполняют несколько функций. Одна из барьерных функций — это изоляция нейронов и их отростков от соприкосновения с кровеносным руслом. Между кровеносными капиллярами и нейронами находятся изолирующие клетки глии. В их функции входят как поддержание целостности гематоэнцефалического барьера, так и питание нейронов. Через эти клетки проходит основной поток веществ и кислорода, необходимого для сохранения активности мозга. Этот глиальный барьер непроницаем для большинства органических соединений. Их перенос к нейронам осуществляется под контролем рецепторных белков мембран глиальных клеток и нейронов. Такой активный фильтр препятствует случайному движению любых соединений как в мозг, так и из него. Через глиальные клетки осуществляется перенос веществ, подвергшихся катаболизму внутри нейронов, поэтому поток соединений через глиальную часть гематоэнцефалического барьера двунаправленный. В мозг поступают кислород и питательные вещества, а из него отводятся продукты катаболизма. Этот поток крайне интенсивен, поскольку у млекопитающих может достигать 25 % общего метаболизма организма. Столь высокий уровень обмена предусматривает высокую проницаемость барьера при невероятно эффективной избирательности. Эти функции структурно обеспечены соотношением количества глиальных клеток и нейронов. Как правило, каждый нейрон обслуживает примерно 15–50 глиальных клеток, которые и обеспечивают необходимый и избирательный поток компонентов, необходимых для поддержания жизни нервной клетки.
Надо отметить, что изолированность нервной системы двунаправленная. Глиальные клетки препятствуют попаданию продуктов, появляющихся при гибели нейронов, и в мозг, и в кровеносную систему. После гибели нейрона такие продукты формируют вокруг него своеобразный саркофаг из своих тел. Это препятствует попаданию продуктов аутолиза в межклеточное пространство. После окончательного распада нейрона остаётся только контур из тел глиальных клеток, формировавших саркофаг, а затем исчезает и он. Появляются своеобразные «тени» — пустые межклеточные участки, напоминающие форму погибших клеток. Гематоэнцефалический барьер мозга построен не только из глиальные клеток. Его функции выполняют и эпендимные клетки, выстилающие поверхность желудочков и сосудистое сплетение (см. рис. I- 10; I-11). Эти клетки в зоне сосудистого сплетения образуют плотный слой, который препятствует проникновению через межклеточное пространство любых веществ и соединений.
Рис. I-11. Срезы мозга и сосудистого сплетения (стрелки), расположенного в желудочках мозга различных позвоночных. Микрофотографии
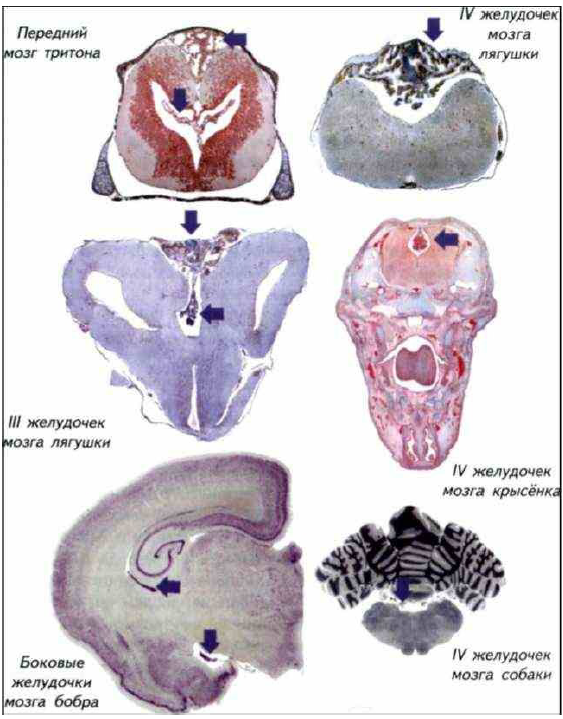
Спинномозговая жидкость фильтруется через эпендимные клетки нейрального происхождения. При низком кровотоке проницаемость стенок сосудистого сплетения невысока, но его площадь очень большая. У млекопитающих при высоком давлении крови сосудистое сплетение имеет крайне небольшие размеры.
Через слой этих клеток в сосудистом сплетении головного мозга происходит ультрафильтрация воды и ионов кальция, натрия, хлора, марганца, калия и магния. Вода и растворы электролитов извлекаются из
плазмы крови. В результате кровь лишается части воды и повышает свою вязкость. Накапливающийся в желудочках фильтрат обычно называют спинномозговой жидкостью. Она проходит через желудочки, стенки мозга и спускается по дорсальной поверхности вдоль спинного мозга, затем поднимается вверх и собирается под мозговыми оболочками в зонах особых расширений. Из них спинномозговая жидкость поступает в специальные зоны мозговых оболочек, которые называются пахионовыми грануляциями. Через грануляции спинномозговая жидкость возвращается в венозное русло. Надо отметить, что спинномозговая жидкость поступает в
головной мозг активно, поскольку артериальное давление в приносящих мозговых сосудах довольно велико, а возвращается в венозное русло уже пассивно — по градиенту концентрации.
Осмотические силы, действующие в момент извлечения спинномозговой жидкости из-под оболочек мозга, не всегда могут уравновесить непрерывный приток этой жидкости через сосудистые сплетения желудочков. Это приводит к динамическим нарушениям и повышению давления жидкости в желудочках мозга. Спинномозговая жидкость меняется в головном мозге с высокой скоростью. У человека, исследованного лучше других животных, при пассивном образе жизни вся вода организма проходит через сосудистое сплетение за 10–12 ч, а при физической нагрузке — за 7 ч. Этот достаточно большой поток жидкости обеспечивает нейроны одним из важнейших факторов жизнедеятельности — растворами электролитов. Они необходимы при кодировке, генерации и передаче электрохимических сигналов между отдельными нервными клетками. Нарушения
электролитного баланса мозга ставят больше проблем, чем недостаток питания нервных клеток. Для контроля за электролитным балансом мозга в эволюции сложилась специальная система, начинающаяся с
осморецепторов, расположенных в прижелудочковых стенках промежуточного мозга. Эти клетки реагируют на изменение осмотического баланса в спинномозговой жидкости. Они вызывают фантомные ощущения сухости во рту, стимулируют выработку антидиуретического гормона,стимулирующего адсорбцию воды в почках, и запускают питьевое поведение.
Возникновение этого сложного механизма автономной регуляции осмотического баланса только подчёркивает его функциональную важность для мозга. В этой системе снабжения мозга растворами электролитов нет никаких прямых контактов между нейронами и клетками иммунной системы. Граница непроницаема для органических
соединений всего организма.
Следует отметить, что у позвоночных сосудистое сплетение различается по размерам (см. рис. I-11). У рыб и амфибий оно выглядит непропорционально большим, а у млекопитающих — чрезвычайно маленьким. В контексте рассуждений о скорости обмена спинномозговой жидкости такие различия кажутся необъяснимыми (Савельев, 2001).
На самом деле причины таких морфологических различий вполне понятны.
Скорость кровотока в сосудистом сплетении у птиц и млекопитающих намного выше, чем у рептилий, амфибий, хрящевых и костистых рыб, поэтому достаточный уровень обмена спинномозговой жидкости у холоднокровных обеспечивается большей площадью поверхности сосудистого сплетения. Отношение площадь поверхности сосудистого сплетения/объём мозга у низших позвоночных в несколько раз больше, чем у птиц или млекопитающих. Известны и «гипертрофированные» исключения из этого правила, например у бурого протоптера (Protopterus annectens) сосудистое сплетение закрывает собой почти всю дорсальную
поверхность мозга.
Таким образом, изолированность и высокий уровень метаболизма нейронов головного мозга обеспечены двумя относительно независимыми системами. Одна из них представляет собой глиальные клетки, обеспечивающие метаболизм питательных веществ и кислорода, другая — эпендимные клетки сосудистого сплетения, фильтрующие через своё тело поток воды и электролитов из плазмы крови. Процессы разделены,
поскольку даже при значительном недостатке пищи электрохимическая активность мозга поддерживается независимо. Это происходит благодаря эффективному и относительно независимому обмену спинномозговой
жидкости и электролитов нервной системы (см. рис I-8; I-10; I-11).
Дополнительное внимание следует уделить изоляции периферической части нервной системы. Она является таким же забарьерным органом, как головной и спинной мозг. Все периферические нервы, ганглии,
рецепторные и эффекторные окончания изолированы от иммунной системы организма. Нервы и ганглии окружает оболочка из особых клеток, которые называются шванновскими (см. рис. I-9; I-10). У позвоночных они
происходят из клеток нервного гребня, как и большая часть периферической нервной системы. Обычный размер этих клеток, окружающих аксоны и дендриты нейронов, составляет около 1 мм.
Шванновские клетки формируют изоляционный слой вокруг отростка нейрона при помощи своей мембраны, которая может образовывать множество витков. В сечении эта структура напоминает плотный рулет
(рис. I-12). В случае особо скоростного проведения сигналов миелинизация может стать «матрёшечной»: внутри общей миелиновой оболочки может лежать высокоскоростной нерв, окружённый собственной многослойной
миелиновой оболочкой. Обычно скорость проведения сигналов по таким нервам более 130 м/с. Зоны контактов отдельных шванновских клеток называются перехватами Ранвье.
Рис. I-12. Оболочки отростков нервных клеток (а, в) и синапсов (б).
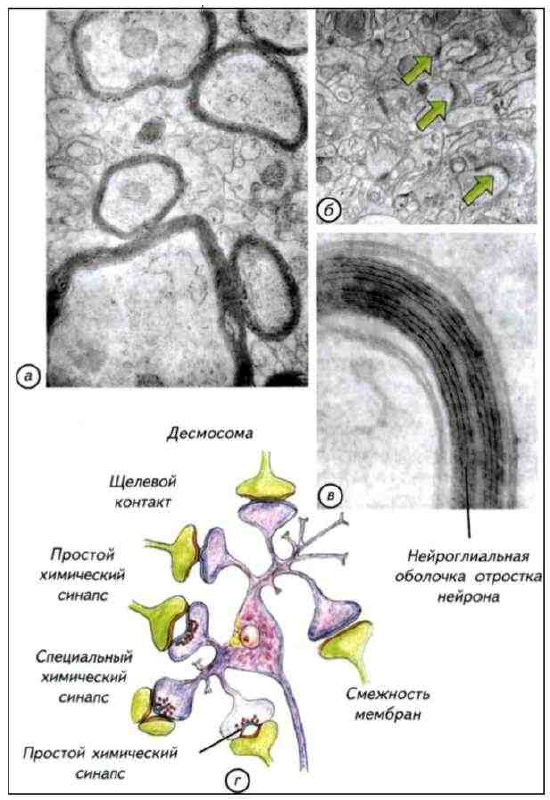
Электронные фотографии. Схема основных типов синаптических контактов нервных клеток (г). Синапсы и контакты увеличены.
Оболочки отростков нервных клеток изолируют зоны проведения сигналов и увеличивают скорость их передачи. Синапсы обозначены зелёными стрелками.
В этих зонах часто располагаются складки мембраны аксонов, которые выходят наружу и формируют эффективно работающие соединения, синапсы. Места контакта нейронов с органами-мишенями также изолированы специализированными гомологами шванновских клеток. Отдельно необходимо пояснить ситуацию с миелинизированными и немиелинизированными (безмиелиновыми) волокнами. Под этим названием обычно понимают волокна, «лишённые» оболочек. Это название укрепилось в учебниках с конца XIX в., но не отражает реальной ситуации. Безмиелиновыми нервными волокнами микроскописты, использовавшие световой микроскоп, считали волокна без явных следов оболочек или миелина. Однако с помощью электронного микроскопа показано, что даже обонятельные нервы обладают небольшой оболочкой, состоящей из шванновских клеток.
Обычно одна шванновская клетка делает 1–2 оборота вокруг группы обонятельных волокон. Тем не менее нервные волокна изолированы на всём протяжении. Вполне понятно, что обновляющиеся обонятельные клетки не могут иметь развитой изолирующей оболочки, хотя в упрощённом виде она всегда присутствует. В периферической нервной системе нет неизолированных участков ганглиев, нейронов или их отростков и концевых разветвлений. Различия сводятся к степени миелинизации, а не к разным принципам строения. Следовательно, в головном и спинном мозге барьерные функции выполняют глиальные клетки, система сосудистых сплетений и олигодендроглия, в периферической нервной системе — шванновские клетки. Нервная система изолирована от остального организма, а нарушение этого барьера приводит к тяжелым аутоиммунным заболеваниям и гибели животного.
Взаимодействия между клетками
Нервные клетки взаимодействуют между собой и с остальными тканями организма. Обычно это прямой контакт. Нервное окончание получает информацию или передаёт её клеткам органа, но это не обязательно. Нервные клетки могут синтезировать гормоны, нейропептиды или другие соединения. Они выделяются в кровеносное русло и распространяются по гуморальным законам. Гормоны используются как генерализованные носители информации для управления всем организмом. Иногда они специфичны только для определённого органа-мишени, но в целом гормональная регуляция очень неспецифична и определяет только общую тенденцию в поведении. Выброс половых гормонов происходит под влиянием нервной системы, но их присутствие в организме в конечном счёте подчиняет себе и работу мозга. Мозг «вызывает их к жизни» и сам подчиняется им. Так, в период гона у копытных стратегически меняется поведенческая активность. Половые гормоны оказывают столь заметное влияние на мозг, что все другие формы поведения отходят на второй план или становятся подчинёнными. Достаточно попробовать плоды блестящей дрессировки любимого домашнего пса в присутствии течной суки.
В человеческом (приматном) сообществе действуют похожие законы. Весенняя гормональная активность преждевременно снимает шапки у мальчиков и оголяет коленки у девочек. Как правило, никакие «негормональные» доводы не действуют. Гормональная подчинённость нервной системы — это интеллектуальное горе человечества и гарантия его воспроизведения как биологического вида.
Размножаться, драться и добывать пищу лучше с использованием гормональной поддержки организма. Древние викинги грызли край щита, доводя до нужного уровень адреналина перед боем. Словесная перепалка на кухне вызывает выброс мобилизирующих гормонов, а через 10 мин становится ясно, как много веских слов и аргументов ещё не высказано. Следовательно, гормональные межклеточные взаимодействия, запускаемые нервной системой, хороши, но инертны, неадаптивны и не поддаются динамическому контролю. Трудно представить, что, собираясь отчаянно спорить, человек будет колоть себя шилом для гормональной мобилизации. Ещё менее вероятен волк, грызущий свой хвост для охотничьего возбуждения.
Для многих других видов гормональный контроль поведения позволяет просто статистически решать проблемы выживания. Для животных с выраженными генетическими программами поведения гормональная регуляция является одним из средств реализации врождённых форм поведения. Это свойственно беспозвоночным, первичноводным позвоночным, амфибиям, значительной части рептилий, птиц и специализированных млекопитающих. Такая распространённость генетико-гормональных форм поведения показывает их эффективность, но основана на вероятностном принципе. У таких видов обычно достаточно много потомков, чтобы хотя бы один из них смог выжить, просто перебирая стандартный набор поведенческих программ.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы