Отношение массы мозга и тела
Мозг первичноводных позвоночных животных (круглоротые, хрящевые и костистые рыбы), амфибий и рептилий составляет примерно 1/10-1/40 массы всей нервной системы. Эта пропорция существенно зависит от размеров тела животного. Если животное крупное, то масса периферической нервной системы, куда входят нервы и ганглии, может более чем в 10 раз превышать размеры головного и спинного мозга вместе взятого. По-видимому, у вымерших крупных динозавров размер головного мозга был крайне мал по сравнению с таковым периферической нервной системы и мог составлять её 30-ю или даже 50-ю часть. У птиц и млекопитающих ситуация несколько иная. У птиц головной и спинной мозг обычно составляет 1/2-1/5 массы всей нервной системы. Существуют и исключения. У мелких птиц пропорция нарушается, поскольку мозг
имеет огромную относительную массу, а тело весит всего несколько граммов.
Рис. 1-6. Отношение масса мозга/масса тела у различных животных.

Мозг/ тело
Колибри 1/12
Мышь 1/26
Крыса 1/28
Крот 1/34
Белка 1/36
Летучая мышь 1/40
Ленивец 1/100
Горилла 1/100
Голубь 1/100
Пчела 1/130
Афалина 1/140
Олень 1/150
Орёл (могильник) 1/160
Шакал 1/200
Собака 1/250
Кролик 1/300
Одиночная оса 1/300
Овца 1/350
Лошадь 1/400
Тапир 1/500
Слон 1/500
Лягушка 1/750
Страус 1/800
Морж 1/850
Лев 1/1000
Тигр 1/1200
Бегемот 1/1500
Жираф 1/1700
Кашалот 1/3500
Жук-плавунец 1/4000
Финвал 1/10000
Синий кит 1/20000
Рис. I-7. Сравнительные размеры спинного и головного мозга у животных различных групп.

Головной и спинной мозг почти равны по массе у лягушки (а); у зелёной мартышки и игрунки масса головного мозга намного превышает массу спинного (6, в). Спинной мозг змеи по размерам и массе во много раз превышает головной мозг (г).
Приматы (мелкие) 1/40
Полёвка (мышь) 1/47
Человек 1/50
Бурозубка 1/50
Домашняя кошка 1/80
Одним из рекордсменов в этой области является колибри. Масса мозга колибри составляет 1/12 массы тела. Для птиц и млекопитающих это рекордное отношение, поскольку оно больше только у ново-рождённого ребёнка (1/7), но новорождённый не может самостоятельно выжить. Колибри считается животным с максимальной относительной массой мозга (см. рис. I-6). Однако в этих схемах не учитываются масса периферической нервной системы и её роль в метаболизме. Если добавить к мозгу колибри ещё и массу её периферической нервной системы, то общее отношение нервная система/тело станет 1/6.
Интересно отметить, что у многих беспозвоночных животных отношение массы мозга к массе тела ненамного отличается от этого показателя у теплокровных позвоночных. Если за головной мозг членистоногих принять головные ганглии с нейрогемальным органом, то его массе могут «позавидовать» многие позвоночные. Так, у рабочей пчелы отношение масса головных ганглиев/масса тела будет 1/130, у муравья — 1/140, у одиночной осы — 1/300, у майского жука и жука-плавунца — 1/4000. Иначе говоря, головные ганглии пчелы и муравья сопоставимы с относительными размерами головного мозга оленя, а одиночной осы — с мозгом льва. Вместе с тем масса туловищных ганглиев беспозвоночных почти всегда превышает массу головного мозга. Если учесть их размеры, то пропорция масса нервной системы/масса тела примет очень неожиданный вид — она достигнет уровня высших млекопитающих. Для пчёл, муравьев, одиночной осы, майского жука и плавунца она составит 1/40,1/45,1/100 и 1/1200 соответственно. Сопоставив эти отношения с аналогичными у приматов, мы получим неприятную «закономерность»: жуки-плавунцы и высшие приматы объединятся в общем «церебральном индексе», что подчеркнёт сомнительную ценность этого параметра для оценки интеллекта. Массовые отношения тела и мозга только косвенно отражают основные тенденции эволюции мозга.
У млекопитающих отношение массы мозга к массе периферической нервной системы имеет ряд особенностей. Головной и спинной мозг в этой группе может превышать массу всей остальной нервной системы. В этом случае отношение мозг/периферическая нервная система может изменяться в пределах от 3/1 до 1/5. Причины таких колебаний скрываются в размерах тела и мозга млекопитающих. Землеройки имеют небольшую массу тела и незначительные размеры периферической нервной системы, отношение мозг/периферическая нервная система будет у них около 3/2.
Однако мозг землероек относительно массы тела очень велик, что принципиально меняет вид пропорции. Общее отношение нервная система/тело с учётом периферической нервной системы станет примерно 1/7.
Совершенно иная ситуация у крупных млекопитающих. Хоботные и китообразные имеют огромный мозг (см. рис. I-6) и огромную массу тела.
Для обслуживания столь крупного тела необходима крайне развитая периферическая иннервация. Приблизительное отношение мозг/ периферическая нервная система приближается к 1/2. Это значит, что у кита-финвала общая масса нервной системы может превышать 20 кг. Для оценки реальных размеров всей нервной системы необходимо учитывать несколько параметров одновременно.
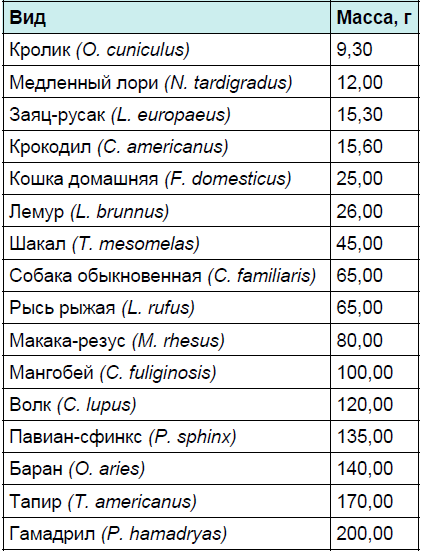
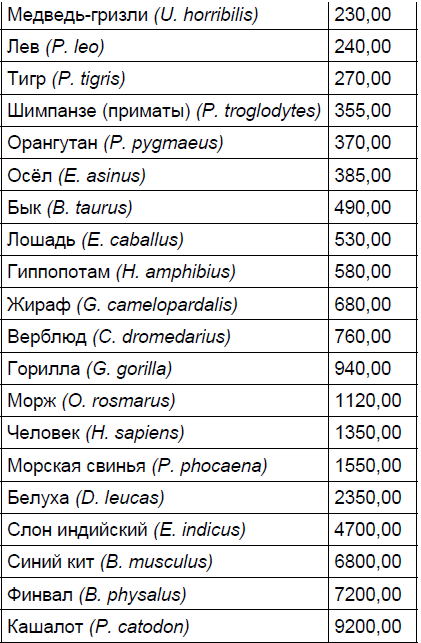
Первым параметром является абсолютная масса мозга, вторым — масса периферической нервной системы, третьим — масса тела животного. Только учёт всех перечисленных параметров даст реальную долю энергетических затрат на содержание мозга. Масса головного мозга различных животных представлена в табл. 1. Из табл. 1 видно, что мозг млекопитающих может различаться по массе в 10 000 раз. Это разнообразие соответствует разнообразию размеров животных. Однако каждый вид весьма неоднороден. Он состоит из подвидов и пород, которые имеют существенные различия как по размерам тела, так и по объёму мозга.
Таблица 1. Абсолютная масса головного мозга у животных разных видов

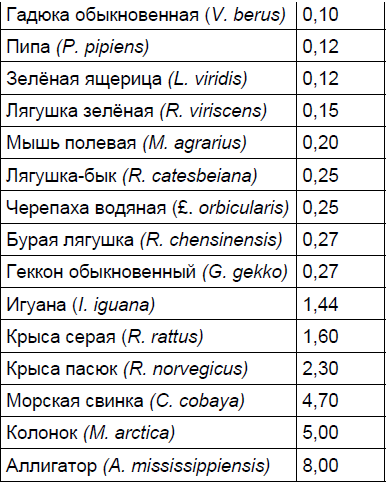
Продолжение таблицы 1
Примером таких различий могут служить собаки. Их мозг может различаться по массе в несколько раз. Самая маленькая масса мозга собаки, которая была описана в литературе, составляла 41,5 г, а самая большая — 212 г (Петропавловский, 1904). Табл. 2 поясняет внутривидовую изменчивость мозга собак.
Таблица 2. Средние величины массы головного мозга породистых и беспородных собак
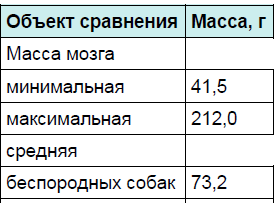

Показательно процентное соотношение массы мозга и массы тела. Минимальное соотношение 0,2 выявлено у крупных собак, масса тела которых составляет 28–44 кг. Для особей с массой меньше 2,5 кг оно увеличивается до 2,9. Следовательно, масса мозга по отношению к единице массы тела уменьшается в соответствии с увеличением размеров животного. Однако у щенков и взрослых собак одной и той же породы отношение массы мозга к массе тела практически одинаково. Это отличает индивидуальное развитие собак от развития мозга обезьян и человека.
У обезьян и человека плод рождается с большим мозгом и маленькой массой тела, а затем масса тела возрастает намного быстрее, чем мозг. Головной мозг подвержен половому диморфизму. У самок мозг обычно на 10–12 г
меньше, чем у самцов. Суки обычно мельче, чем кобели, а их относительная масса мозга больше.
Весьма любопытны различия в этом плане породистых и беспородных собак. Средняя масса мозга в выборках беспородных собак колеблется от 42 до 118 г, а породистых — от 41,5 до 212 г. Породистые животные имеют большую амплитуду изменчивости головного мозга, чем беспородные. По- видимому, у одичавшей в Австралии собаки Динго мозг ещё более однороден по массе, чем у беспородных городских собак. Возврат к природным формам уменьшает искусственно созданное разнообразие.
Надо отметить, что у породистых собак относительная масса мозга почти в 2 раза меньше, чем у беспородных.
Существует индивидуальная изменчивость размеров мозга у различных видов в природе. В специальных исследованиях на 25 зайцах-русаках (Lepus europaeus), проведённых в Польше, показана существенная
изменчивость мозга. При средней массе мозга 15,3 г были найдены особи с мозгом массой 12 и 17 г. Известны вариации в массе мозга у приматов, хищных, копытных и сумчатых; они обычно не превышают 2024 %.
Сходная изменчивость обнаружена и в массе мозга птиц, которые обитают в Европе и Америке (Lefebvre ef al., 1997).
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы