Теория переходных сред
В основе эволюции нервной системы позвоночных лежат общие морфологические закономерности. Они сводятся к количественным и качественным изменениям в центральной и периферической нервной системе. Однако в отличие от других систем организма, любые структурные изменения вызывают глубокую перестройку поведения.
Результатом становится изменение форм взаимодействия организма с внешней средой. Новые морфофункциональные свойства нервной системы не всегда приводят к положительным результатам. Одни из этих свойств становились основой для кратковременного процветания группы или тупиковой специализации, другие давали позвоночным возможность освоить бескрайние ресурсы и открывали перспективные пути эволюции. В естественной истории нервной системы были и остаются морфологические решения, обрекающие своих владельцев как на неизбежное вымирание, так и на процветание. Большинство современных животных представляют собой образцы более или менее удачных, но тупиковых адаптаций. Их исчезновение было предопределено в момент начала структурной специализации нервной системы.
Нервная система обладает одним примечательным свойством: он может практически мгновенно изменить поведение животного и физиологическую активность его органов, а затем столь же быстро восстановить исходную ситуацию. Обратимость быстротечных и абсолютно необходимых изменений делает её бесценным инструментом в биологическом мире. Однако диапазон возможных перестроек нервной системы ограничен её структурой. Мозг может предоставить только тот набор инстинктивных или ассоциативных решений, который обеспечен сенсомоторными системами организма. Медведь не станет махать лапами, даже если реально сможет взлететь. На такие поступки легко решается только цивилизованный человек, поскольку его мозг почти утратил связь с реальным миром планеты. Иначе говоря, все позвоночные животные оказываются пленниками эволюционного прошлого своей нервной системы. Позволяя животным быстро приспосабливаться к сиюминутным изменениям окружающей среды, мозг формирует своеобразные скрытые рамки предельно возможных изменений. Именно эти рамки и предопределяют пределы обратимых адаптивных изменений поведения конкретного вида.
Морфологическая эволюция нервной системы необходима как инструмент по расширению границ адаптивных возможностей.
Структурные изменения мозга снимают ограничения с одних поведенческих реакций и формируют другие. Этот процесс может продолжаться до тех пор, пока не возникнет слишком специализированный для дальнейших перестроек мозг. Тем не менее количественные или качественные изменения нервной системы остаются единственным выходом за пределы стандартных возможностей. Надо подчеркнуть, что
количественные изменения в нервной системе могут происходить намного быстрее, чем качественные. Именно они являются первичным ресурсом для структурных адаптаций нервной ткани. Качественные морфологические перестройки мозга крайне затруднены и обычно требуют специальных условий или продолжительного времени. Эта разница между количественными и качественными изменениями нейральных структур опосредована особым положением головного и спинного мозга в системе органов и тканей позвоночных.
Центральная нервная система не участвует в биомеханических взаимодействиях организма с внешней средой. Это не значит, что при сокращении мускулатуры, движении суставов, деформациях кожи или при движении пищи по кишечнику нервы не подвергаются механическим нагрузкам. Они обладают известной прочностью и пластичностью и могут выдерживать небольшие и краткосрочные нагрузки. Однако речь идёт не о механических свойствах нервной системы. Наоборот, для эволюционной морфологии наиболее интересно, что нервная система предельно защищена от любых нагрузок, за исключением специализированных механорецепторов. Вся эволюция головного и спинного мозга проходит внутри черепа и под защитой невральных дужек позвонков. От элементов скелета они отделены тремя мозговыми оболочками и спинномозговой жидкостью. Однако форма центральной нервной системы незначительно зависит от скелета. Достаточно упомянуть, что в эмбриональном периоде развития головной и спинной мозг является индуктором дифференцировки скелета, а не наоборот. Справедливее сказать о том, что форма черепа и невральных дужек позвонков вторична по отношению к анатомии центральной нервной системы. Следовательно, головной и спинной мозг изменяет свою форму независимо от каких-либо биомеханических преобразований в организме животного.
В этом особом положении мозга заключён огромный потенциал любых структурных преобразований. Реальным источником количественных изменений является стабильная индивидуальная изменчивость нервной системы. Специальные исследования, проведённые на амфибиях, рептилиях и млекопитающих, показали, что у жизнеспособных личинок или молодых животных, полученных от одной пары родителей,
стандартной является 20–22 % количественная изменчивость головногомозга. Нейроны подсчитывали как во всех отделах мозга, так и в основных периферических анализаторах. Был обнаружен некоторый разброс значений изменчивости в зависимости от отдела головного мозга.
Наиболее древним структурам (задний и продолговатый мозг) была свойственна 7-13 % изменчивость, а для эволюционно новых — 18–25 %.
Тем не менее количественной вариабельностью были охвачены практически все отделы головного мозга. Выявленные пределы изменчивости установлены в генетически однородной группе животных — потомков только одной пары родителей.
Если использовать человека для оценки изменчивости нервной системы, то двукратная разница по массе мозга далеко не исчерпает все встречающиеся и жизнеспособные варианты (Савельев, 1996), поэтому наиболее объективным будет вывод о нормальной внутривидовой 15–25 % изменчивости головного мозга. Это означает постоянное отличие одного животного от другого на некоторое количество нервной ткани. Для анамний этот ресурс может составлять от нескольких тысяч до десятков миллионов нейронов, а для амниот от сотен тысяч до нескольких миллиардов клеток. Учитывая, что каждый нейрон обладает множеством контактов с другими клетками и может быть носителем памяти, мы можем предполагать заметную разницу в поведении отдельных особей даже в самой однородной популяции. Этологические подтверждения
индивидуализации поведения многочисленны и охватывают практически все группы позвоночных. Это означает, что в любой популяции существуют особи, которые могут решать определённые задачи лучше или хуже, чем другие. Если биологическая ситуация стабильна, то указанной разницей в потенциальных возможностях мозга никто и никогда не воспользуется.
Количественные различия мозга становятся значимыми при нестабильности окружающей среды, высокой половой конкуренции или явном, но малодоступном пищевом ресурсе. Они оказываются решающим резервом при полной исчерпанности инстинктивно-ассоциативных наборов видоспецифических форм поведения. Если индивидуализированная форма поведения даёт заметный выигрыш в доступе к пище, то она и закрепляется последующими репродуктивными преимуществами, вероятность сохранения количественных особенностей мозга данной особи возрастает.
По-видимому, именно этот механизм лежит в основе масштабных приспособительных изменений головного мозга большинства первичноводных позвоночных. В зависимости от типа питания и развития органов чувств их мозг дифференциально увеличивается в размерах (см. § 27). Этот путь в эволюции нервной системы эффективен для решения частных адаптивных проблем в рамках существующей конструкции нервной системы. Крупные эволюционные события, приводящие к смене среды обитания и возникновению новых систематических таксонов высоких порядков, требуют качественных изменений в нервной системе.
Появление в нервной системе качественно новых структур требует продолжительного времени и совершенно особых условий. Эти условия должны отличаться от традиционной среды обитания и обладать непреодолимой притягательностью для позвоночных. Гарантиями такой привлекательности остаются обильная пища и успешное размножение.
Если столь биологически выгодная среда долго сохраняется, то у животных появляется шанс обретения качественно новой нейроморфологической структуры.
Таких экологических условий в истории позвоночных возникало немного, и все они отмечены появлением животных с качественно новыми структурами нервной системы. Первым событием такого рода стало возникновение хордовых. Как описано выше, появление хордовых было довольно случайным событием, а не фатальной эволюционной закономерностью (см. § 26). Группа похожих на турбеллярий небольших
плоских червей продолжала обитать на мелководьях, богатых пищей.
Будучи фильтраторами и ведя пассивный образ жизни, эти плоские червеобразные существа пытались закрепиться на максимально выгодных пищевых территориях. Для этого они погрузили заднюю часть своего тела в придонные отложения. Такое заякоривание широко распространено среди современных придонных беспозвоночных. Отдалёнными последствиями этих несложных адаптивных действий древних червей стали дорсальный нервный тяж и мышечная хорда, предотвращающая его деформацию. Сутью качественных изменений двух- или четырёхцепочечной нервной системы червеобразных предков хордовых стали несколько последовательных событий. При двухцепочечном варианте произошёл 90-градусный поворот червя на одну из боковых поверхностей тела. При четырёхцепочечной схеме строения нервной системы отмечено слияние парных дорсальных и вентральных нервных цепочек. В обоих случаях качественная перестройка нервной системы завершилась рострокаудальным слиянием сегментарных ганглиевдорсальной нервной цепочки с последующим образованием центрального желудочка.
Параллельно произошло разделение узлов вентральной нервной цепочки до уровня соматических ганглиев (см. § 26). Они стали основой для иннервации внутренних органов. Хордовые не появились бы без специфической переходной среды. Небольшая глубина воды, обилие пищи и подходящие для размножения условия гарантировали процветание любых придонных фильтраторов. Среди множества вариантов адаптации к столь благоприятной среде возникновение морфотипа хордовых было только одним из успешных вариантов. В этой ситуации решающую роль играла богатая пищей среда, которая стала стимулом для морфологических изменений многих видов. Дальнейшая эволюция хордовых протекала в более разнообразных условиях и привела к возникновению всего многообразия первичноводных позвоночных (см. § 29).
Вторым принципиальным качественным изменениям мозг подвергся после выхода позвоночных на сушу.
Это событие привело к крупным морфологическим перестройкам как в нервной системе, так и других органах. Сформировались конечности, лёгочное дыхание, специализированные покровы и ряд других признаков, позволивших архаичным тетраподам перейти к наземному существованию. Столь обширные морфофункциональные перестройки анализаторного и эффекторного аппаратов нервной системы не могли произойти за короткий промежуток времени и вне особой переходной среды. Они были особенно необходимы для качественных изменений в нервной системе, поскольку в количественном отношении мозг амфибий явно проигрывает специализированным первичноводным животным. При выходе на сушу в нервной системе древних амфибий возникли вомероназальная обонятельная система, контроль за дыханием и комплекс стволовых центров управления конечностями. Изменениям подверглась зрительная, слуховая и вестибулярная системы. Переходной экосистемой между водной и наземной средой обитания могли быть своеобразные почвенные лабиринты или карбоновые лесные завалы (см. § 31). В такой переходной среде можно было долго использовать как плавательные движения, так и опору на плавники.
При высокой влажности лабиринтов одновременно функционировали кожное дыхание, жабры и зачатки лёгких. Развитие водно-воздушных органов чувств и моторных систем в переходной среде было оправдано биологическими преимуществами, которые давало освоение богатых пищей и хорошо защищённых территорий (см. § 33). По- видимому, и почвенные лабиринты, и карбоновые завалы из стволов деревьев создали уникальную переходную среду для постепенной эволюции нервной системы древних амфибий. Только при длительном развитии морфологических изменений могли бы появиться спинномозговые центры и красное ядро для управления конечностями, вомероназальный орган и дополнительная обонятельная луковица, вторичные слуховые и вестибулярные центры.
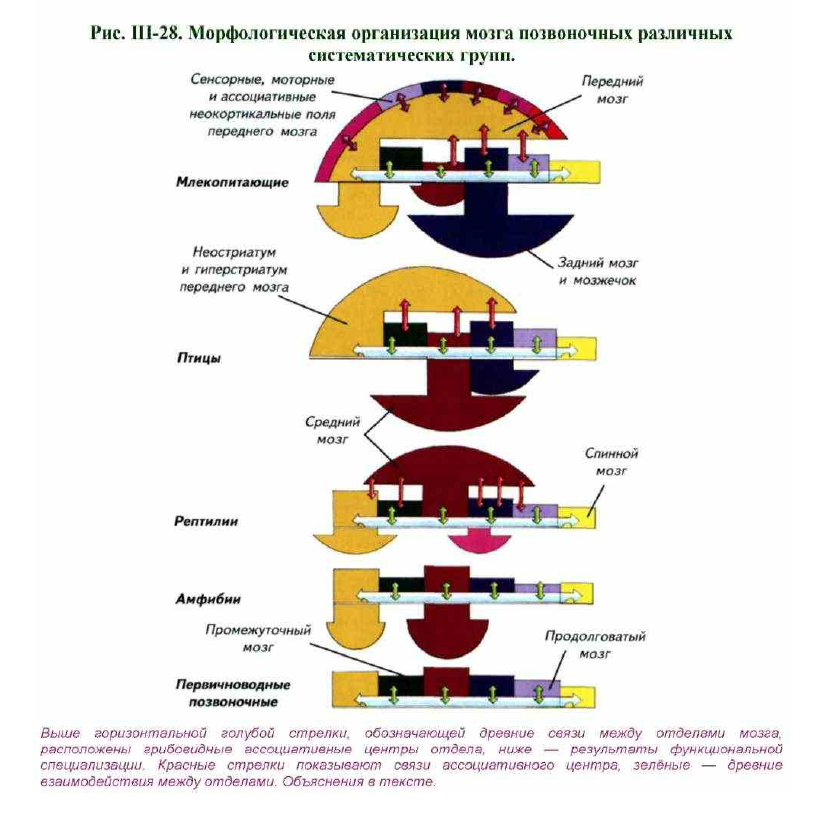
Третьим историческим периодом развития нервной системы можно считать формирование мозга архаичных рептилий. Рептилийный период стал самым плодотворным в истории позвоночных. Рептилии заложили основные принципы структурной эволюции мозга амниот. У рептилий в нервной системе впервые сформировался ассоциативный отдел. Он возник на базе среднего мозга и оказался настолько успешным приобретением, что рептилии на миллионы лет стали самой доминирующей группой позвоночных. Ассоциативный средне-мозговой центр никогда не сформировался бы без серьёзной биологической необходимости. Она возникла ещё в начале эволюции рептилий как способ адаптации к агрессивной среде. Архаичным рептилиям требовалось постоянно сравнивать информацию, приходящую от различных органов чувств, и принимать сложные решения. Решения были вызваны постоянной адаптацией поведения к быстро меняющейся ситуации. Этими свойствами мозг первичноводных позвоночных и амфибий не обладал. Они выбирали одну из инстинктивных форм поведения по совершенно другим принципам. Выбор амфибий был построен на конкуренции между мозговыми центрами представительства анализаторов (рис. III-28).
Простое сравнение уровня возбуждений было достаточным условием дляреализации одной из инстинктивных программ. Рептилии впервые стали обладателями аналитического устройства совершенно нового типа (см. рис.III-28).
Оно действовало по принципу сравнения информации,поступающей от каждого органа чувств. Решающую роль стало играть содержание анализаторного сигнала, а не сам факт возбуждения (см. § 37).
Собственно говоря, у рептилий появились основы ассоциативного принципа поиска решений. Понятно, что мы видим самые зачаточные признаки этого губительного свойства мозга, но они возникли именно у рептилий. История рептилий, наверное, была намного богаче неврологическими экспериментами, чем мы можем себе представить.
Достаточно упоминания о ещё одном историческом приобретении рептилий — кортикальных структурах переднего мозга (см. § 39). Половая конкуренция в сочетании с невероятным развитием обоняния и вомероназальной системы рептилий стала основой для появления кортикальных структур. Кортикальные структуры переднего мозга сформировались на основе нового центра, обеспечивающего интеграцию
половых сигналов с остальными органами чувств. Этот половой интегративный центр непродолжительное время конкурировал с ассоциативной крышей среднего мозга, но его активность проявлялась только в период размножения. По-видимому, для успешного размножения архаичным рептилиям нужно было подчинять все системы организма этой задаче, а любые побочные занятия вплоть до поиска пищи должны были игнорироваться (рис. III-29).
Ассоциативные и кортикальные центры мозга рептилий не могли бы появиться без весьма своеобразных условий. Однако допустим, что архаичные рептилии просто расселились по поверхности Земли. Без серьёзной конкуренции со стороны амфибий, насекомых и растений они быстро стали бы доминирующей группой без глубоких перестроек нервной системы. В таких условиях для её совершенствования нельзя представить никаких реальных оснований. Тем более невозможно отыскать внешние причины для формирования настолько гипертрофированного обоняния, что это привело к возникновению кортикальных структур переднего мозга.
Следовательно, реальные события развивались по совершенно иному сценарию и к идиллическому разбреданию рептилий по поверхности планеты отношения не имели.
Наиболее вероятна довольно длительная эволюция архаичных рептилий в специализированной переходной среде. Эта экологическая ниша, очевидно, была не приспособлена для безмятежного процветания молодой группы позвоночных. Скорее всего все неврологические приобретения рептилий возникли как адаптивные приспособления к крайне сложной среде обитания и агрессивному конкурентному окружению. Такой средой вполне могли быть карбоновые древесные завалы из стволов растений (см. § 38). Эту среду отчасти использовали ещё амфибии, но они явно пришли туда за обильной и гарантированной пищей. Пищей скорее всего были первичноводные позвоночные, которые использовали карбоновые завалы в качестве удобных мест для размножения. Со временем они сменили места размножения или отступила вода. Когда источник пищи по тем или иным причинам иссяк, амфибии стали использовать в пищу себе подобных. Это привело к невиданной конкуренции и быстрому отбору по свойствам и ассоциативным возможностям мозга.
Переходной средой для формирования рептилий стали карбоновые растительные завалы, где трёхмерная среда предъявляла повышенные требования к вестибулярной системе и дистантным анализаторам. Отсутствие света выводило обоняние на качественно иной уровень морфофункционального развития. Он использовался как важнейший дистантный анализатор и система контроля полового поведения. Активно эволюционировала слуховая система, которая не менее эффективна для ориентации в темноте.
За несколько десятков миллионов лет жесточайшей конкуренции в карбоновых растительных лабиринтах сложился уникальный рептилийный мозг с довольно совершенным набором неврологических структур и эффективным ассоциативным центром. С его помощью решались проблемы поиска пищи, конкуренции, избегания опасности и др. Когда наступал период размножения, весь мозг подчинялся новой корковой структуре в стенке переднего мозга. Она стала специализированным центром управления половым поведением, которого не было ни у кого из позвоночных до рептилий. Таким образом, мозг архаичных рептилий стал совершеннейшей системой для решения самых главных биологических задач любого вида — выживания и размножения. Для каждой задачи появилась собственная интегративная система, которая в состоянии перенацелить весь организм рептилий на её решение. С таким поведенческим ресурсом рептилии вышли из своей агрессивной колыбели и очень быстро стали доминирующей группой на планете.
Возникновение мозга птиц нельзя считать принципиальным эволюционным событием, связанным с качественной перестройкой мозга.
Птицы, вероятно, должны были исчезнуть вскоре после своего появления.
Это была тупиковая адаптивная специализация, которую спасла утрата обоняния. Огромный неврологический субстрат обонятельной системы достался архаичным птицам из-за смены пищевых пристрастий. Перейдя к питанию в мелководных заводях или с плавника, они перестали использовать обоняние в качестве ведущей системы афферентации.
Основной анализаторной системой стало зрение, а дополнительной — слух (см. § 43). Добывая пищу в воде, архаичные птицы передвигаться на задних конечностях, что постепенно привело к значительному снижению нагрузки на передние конечности и частичной рудиментации кисти. Роль переходной среды в этом случае играло богатое пищей прибрежное мелководье, которое сохранило притягательность для птиц до настоящего времени.
Хотя узкая специализация птиц гарантировала им быстрое вымирание, переход к плаванию и нырянию за пищей привёл к развитию крылоподобных передних конечностей. На этом этапе эволюции птиц, по-видимому, появились пингвины, которые никогда не летали. Ныряние и плавание с использованием передних конечностей создали физические условия для развития полых костей, мощных грудных мышц, системы воздушных мешков лёгких и перьевого покрова. Судя по всему, добывание пищи в холодных водах стало одним из основных стимулов к обретению теплокровности. Крылоподобные плавательные конечности использовались не только для плавания. Древние птицы применили машущие движения передних конечностей для своеобразного «бега по воде», который стал переходной фазой к активному полёту (см. § 44).

Крылья и перьевой покров сформировались для охоты в водной среде, но были адаптированы и использованы для полёта. В этой ситуации переходной средой стала вода. Она создала все необходимые условия для постепенного накопления изменений в нервной системе птиц, поэтому появление крыльев и переход к полёту не вызвали радикальных перестроек в центральной нервной системе (см. § 43). В связи с редукцией обоняния у птиц на основе базальных структур переднего мозга сформировались ассоциативные центры. Эти центры представлены нео- и гиперстриатумом, которые стали основой становления сложного поведения птиц, памяти и индивидуализации поведения.
Млекопитающие представляют собой довольно странную в неврологическом отношении группу. Преимущества их мозга возникли на основе развития интегративных функций половой системы. Как сказано выше, основной причиной появления кортикальных структур мозга рептилий было развитие вомероназального (якобсонова) органа. Его центральное представительство сформировалось вне древних обонятельных ядер переднего мозга. Основными вторичными центрами вомероназального обоняния стали скромные кортикальные структуры рептилий (см. § 39). На этом морфологическом субстрате сложилась интеграция полового поведения всего организма рептилий. Такое централизованное управление позволяло подчинять весь организм одной задаче и эффективнее достигать успеха в размножении.
Млекопитающие пошли намного дальше рептилий. На этой репродуктивно-интегративной морфологической структуре переднего мозга сформировался ассоциативный центр совершенно нового типа. Он стал выполнять функции контроля за работой уже сложившихся сенсорных систем. Автономные механизмы мозга остались на уровне древних центров, а все сложные благоприобретённые функции складывались на уровне коры переднего мозга. Кроме обоняния и половых интегративных центров, для мозга млекопитающих характерно развитие сенсомоторной системы и механизмов кинестетического контроля. Только у млекопитающих мозжечок сформировал парные полушария. Он достиг столь гигантских размеров, что его поверхность зачастую превосходит размеры неокортекса. Более того, значительная, а иногда и большая, часть самого неокортекса обеспечивает соматические, сенсомоторные и моторные функции.
Для появления столь странной специализации нужна весьма оригинальная среда. Сами карбоновые растительные завалы были сложной трёхмерной средой для рептилий, но их мозжечок не достиг даже развития мозжечка птиц. Переходная среда возникновения млекопитающих должна была предъявить необычно высокие требования к анализу положения тела и координации движений. На поверхности земли только в ветвях деревьев могут оказаться столь жёсткие требования к кинестетическому контролю.
По-видимому, в кронах деревьев сформировались все основные сенсомоторные, обонятельные и слуховые преимущества млекопитающих.
Этой переходной средой можно объяснить как появление неокортекса, таки развитие соматической чувствительности, которая стала одним из основных органов чувств (см. § 48).
Результатом становления соматической чувствительности стали рецепторные образования дермы — волосы. Волосы, иннервированные свободными нервными окончаниями, эффективно повысили соматическую чувствительность и затем стали причиной появления волосяного покрова.
Дальнейшее использование волос для терморегуляции замаскировало их первичное предназначение. В кронах деревьев впервые возникло совершенно новое требование к нервной системе (см. § 49). Для архаичных древесных млекопитающих было недостаточно сравнительного анализа информации, поступающей от различных органов чувств. Этот способ работы ассоциативных систем не позволял сделать прогноза событий. В кронах деревьев предвидение развития событий стало решающим условием как для добывания пищи, так и для элементарного сохранения жизни. Только полёт мог бы избавить млекопитающих от этих проблем.
Однако к нему прибегли лишь рукокрылые после формирования основных принципов строения мозга млекопитающих. Основными структурными последствиями обитания в кронах деревьев стали неокортекс, двухполушарный мозжечок и небольшая способность прогнозирования развития событий. Эта особенность млекопитающих после их переселения на почву и в водную среду создала им значительные поведенческие преимущества. Способность к оценке возможных событий стала для млекопитающих инструментом доминирования на планете.
Все перечисленные глубокие изменения в строении нервной системы позвоночных вызваны приспособлением мозга к обитанию животных в специфической среде. Без длительно существующей переходной среды не будет достаточного времени для изменения структурной организации нервной системы. Она слишком качественно консервативна и количественно пластична для быстрых и радикальных морфологических преобразований. Предположение о существовании переходных сред может объяснить причины возникновения мозга современных позвоночных.
С.В.Савельев. Происхождение мозга. Предисловие
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы