27. Мозг первичноводных позвоночных
Рассмотрим основные принципы строения универсальной конструкции нервной системы первичноводных позвоночных (рис. II-17; II-18). Её морфологической осью является центральная нервная система, которая расположена над позвоночником (рис. II-19, а) и включает в себя два отдела: головной и спинной мозг (см. рис. II-17, а). С головным и спинным мозгом соединены периферические нервы. Они проходят, переключаясь
или не переключаясь, через ганглии и связывают периферические органы с центральной нервной системой.
Периферические нервы неодинаковы; часть из них двигательные, или эффекторные. Они передают сигналы из нервной системы к органам-мишеням. Такими органами могут быть поперечнополосатая или гладкая мускулатура, эндокринные железы или секреторные клетки. У ланцетника сами мышечные клетки образовывали отростки, соединяющие их с нервной трубкой. У остальных позвоночных ситуация обратная: двигательные отростки нейронов выходят из мозга или ганглиев, достигают мышц и оканчиваются на поверхности волокон, образуя нервно-мышечные синапсы.
Другая часть периферических нервов чувствительная. Они, наоборот, передают сигналы от внутренних органов и дистантных анализаторов в головной и спинной мозг. Концевые чувствительные участки рецепторных нервов очень сильно различаются по морфологическому строению и рецепторным возможностям. Это позволяет получать дифференцированную информацию различного типа. Особую группу чувствительных клеток и волокон представляют собой дистантные рецепторные органы, или внешние органы чувств. У первичноводных позвоночных к внешним органам чувств относят обоняние, зрение, вкусовые рецепторы, органы боковой линии, рецепторы углового и линейного (гравитационного) ускорения, электрорецепторы и осязание.
Внешние органы чувств сосредоточены в головной части позвоночных, а их нервы приходят в головной мозг. В головной мозг приходит и большая часть информации от внутренних органов, мускулатуры и поверхности тела. По сути дела головной мозг является скоплением клеток, обрабатывающих информацию, поступающую от органов чувств. В соответствии с этими функциями головной мозг имеет специализированные центры, которые обслуживают несколько внешних или внутренних органов чувств. Такие центры по традиции называют отделами мозга.
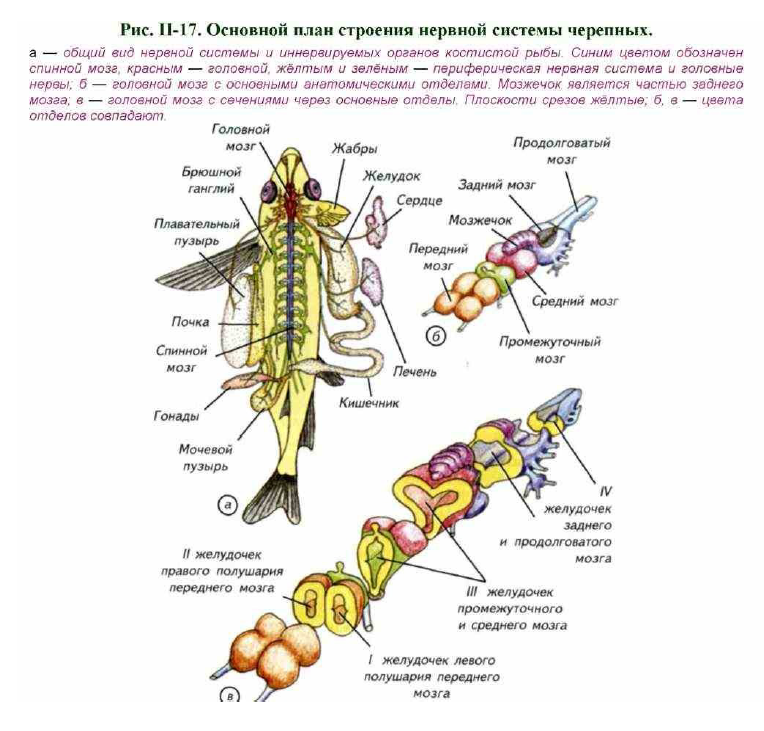
В головном мозге всех позвоночных выделяют 5 специализированных отделов: передний мозг (telencephalon), промежуточный (diencephalon), средний (mesencephalon), задний мозг с мозжечком (metencephalon, cerebellum) и продолговатый мозг (metencephalon) (см. рис. II-17, б). У первичноводных позвоночных отсутствуют полушария мозжечка, а следовательно, и мост, поэтому определить анатомическую границу между
задним и продолговатым мозгом невозможно. Это приводит к тому, что во многих сравнительно-анатомических работах название «задний» и «продолговатый» мозг не применяется. Вместо них используется более общее название — «ромбэнцефалон» (rhombencephalon), которое включает в себя оба отдела.
Понятно, что отсутствие преемственности в названиях не сказывается на гомологизацию этих участков мозга как у первичноводных позвоночных, так и у млекопитающих. Гомология отделов головного мозга сохраняется во всех группах позвоночных (см. рис II-18). Надо отдать должное неординарному сравнительному нейроанатому H.H. Миклухо- Маклаю. Ещё в 70-х годах XIX в. он провёл детальные исследования
головного мозга различных первичноводных позвоночных и их эмбрионов.
Он показал, что основные отделы мозга позвоночных гомологичны друг другу и, несмотря на все эволюционные специализации, сохраняют общность морфологического строения. Результатом многолетних трудов H.H. Миклухо-Маклая стала первая обоснованная гомологизация отделов мозга позвоночных (Миклухо-Маклай, 1952). Уже во 2-м издании сравнительной анатомии К. Гегенбаур (Gegenbaur, 1898) воспользовался этой работой и ввёл представление о нейрогомологиях отделов головного мозга в научный обиход.
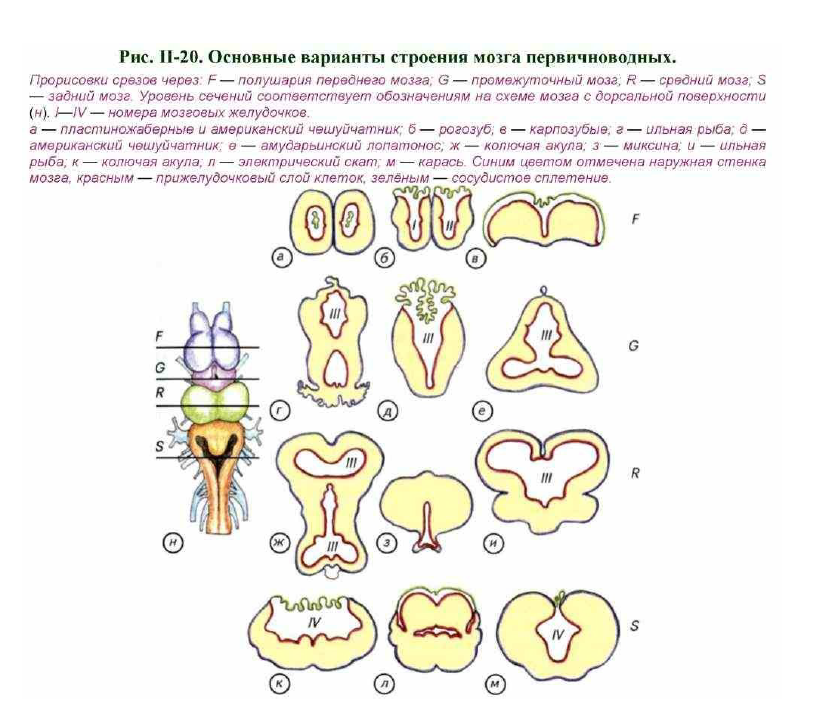
Проводя морфологический анализ мозга первичноводных позвоночных, H.H. Миклухо-Маклай показал, что при изучении отделов мозга необходимо опираться как на внешнее строение, так и на морфологию внутренних полостей — мозговых желудочков. Они имеют специальные названия в каждом из отделов. В парных полушариях переднего мозга находятся первый (I) и второй (II) латеральные желудочки (см. рис. II-17, в; рис. II-20, a-в). Эти два желудочка соединены между собой межжелудочковым отверстием. Они переходят в третий (III) желудочек, который лежит внутри промежуточного и среднего мозга. В латеральных желудочках расположено непрерывное переднее сосудистое сплетение, которое распространяется примерно на 1/2 III желудочка (см. рис. II-20, а- в, д). В свою очередь он переходит в четвёртый (IV) желудочек, а последний продолжается в центральный канал спинного мозга (см. рис. II- 17, б, в). Сверху IV желудочек прикрыт монослоем клеток и содержит заднее сосудистое сплетение (см. рис. II-20, к-м).

Важнейшую часть центральной нервной системы первичноводных позвоночных представляет спинной мозг. Граница между спинным и головным мозгом достаточно условна, поскольку отростки клеток из головного мозга проникают в спинной и наоборот (Nieuwenhuys, 1998).
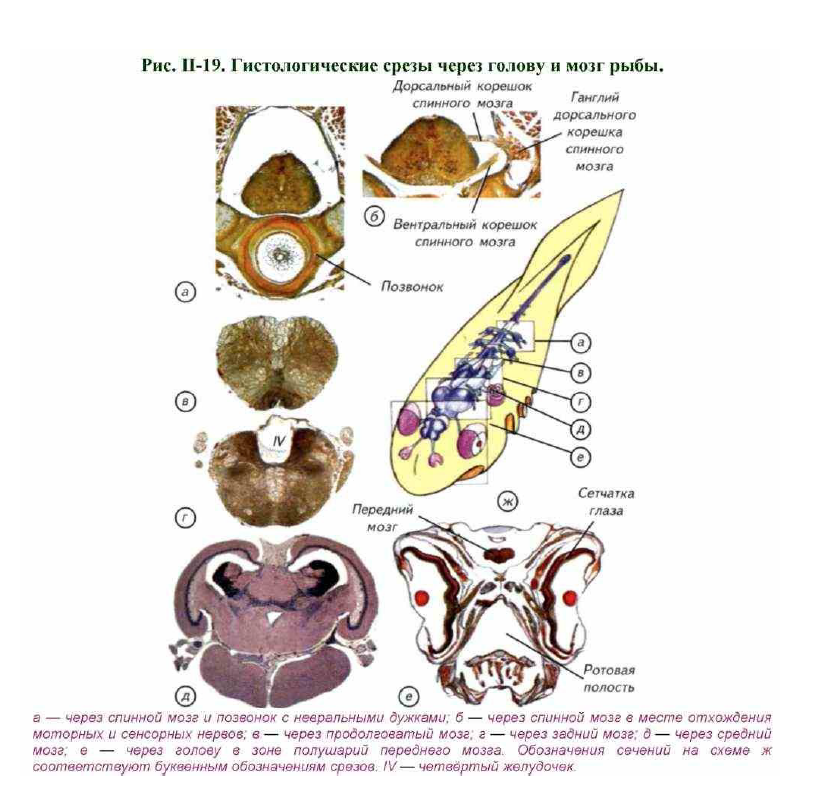
Головной мозг, как правило, расположен дорсально и окружён хрящами или костями черепной коробки. Спинной мозг лежит в полости позвоночного канала, который образован невральными дугами позвонков (см. рис. II-19, а).
У большинства позвоночных спинной мозг имеет однотипную гистологическую структуру. Тела нервных клеток (серое вещество) обычно расположены вокруг центрального канала в виде классической «бабочки», свойственной большинству амниот. У анамний картина несколько смазана, и расположение тел нейронов на разрезах спинного мозга в виде «бабочки» обычно не встречается (см. рис. II-19, а, б). Верхнюю часть серого
вещества называют дорсальными (спинными) чувствующими рогами, а нижнюю — вентральными (брюшными) двигательными рогами спинного мозга. Спинные рога обычно содержат мелкие вставочные нейроны, а брюшные — крупные моторные. Через спинномозговые нервы осуществляются соматическая и висцеральная чувствительность, проходит соматически-двигательная и висцерально-двигательная иннервация.
Соматическая чувствительность включает в себя рецепцию кожных, сухожильных, связочных и мышечных сигналов. Висцеральная чувствительность включает вкусовую рецепцию и сигналы от внутренних органов. Соматически-двигательная иннервация обслуживает скелетные мышцы, а висцерально-двигательная — железы, глоточную, лицевую мимическую и челюстную мускулатуру, гладкую мускулатуру кожи, сосудов и внутренних органов. В состав спинного мозга традиционно включают ганглий дорсального корешка спинного мозга, или спинальный ганглий, в котором расположены тела афферентных (чувствительных) соматических и висцеральных нейронов (см. рис. II-19, а, б). Эти клетки связаны со спинным мозгом коротким отростком, который оканчивается в дорсальных рогах серого вещества. Вентральная часть спинного мозга составлена из эфферентных (двигательных) волокон. Они начинаются от клеток, расположенных в вентральных рогах спинного мозга (Савельев, 2001).
Не у всех позвоночных есть чёткое разделение корешков спинного мозга на моторные и сенсорные. У многих первичноводных позвоночных и амфибий висцеральные эфферентные волокна выходят из спинного мозга как через дорсальные, так и через вентральные корешки.
У ланцетника, миног и миксин висцеральные эфферентные волокна преимущественно выходят через дорсальный корешок спинного мозга.
Надо отметить, что слияния дорсальных и вентральных корешков спинного мозга у ланцетника и круглоротых не происходит. Они в виде самостоятельных нервов достигают иннервируемых органов. У большинства высших позвоночных дорсальные и вентральные корешки спинного мозга выходят на одном уровне. У ланцетника, миног, миксин и акул дорсальные корешки спинного мозга чередуются с вентральными.
Они лежат на разных уровнях, обозначая границы миотомов. Эфферентный соматический (моторный) корешок выходит из спинного мозга в центре миотома, а афферентные (чувствительные) и висцеральный эфферентный — в промежутках между миомерами. Таким образом, наиболее примитивными характеристиками морфологической организации спинальных нервов можно считать чередование дорсальных и вентральных корешков, их самостоятельность и висцеральные эфферентные волокна в дорсальных корешках спинного мозга.
Спинной мозг выполняет ряд автономных функций и интегрирует свою активность с головным мозгом. Эффективность этой интеграции определяется нервными связями, которые организованы в виде восходящих и нисходящих путей спинного мозга. Восходящие и нисходящие волокна спинного мозга организованы таким образом, что восходящие локализуются преимущественно в дорсальной части спинного мозга, а нисходящие в вентральной. Восходящие пучки волокон направляются в головной мозг и оканчиваются в 5 основных центрах: мозжечке, моторных центрах заднего и продолговатого мозга, промежуточном мозге и крыше среднего мозга. Нисходящие волокна расположены преимущественно в вентральной половине спинного мозга. Основная часть нисходящих волокон спинного мозга первичноводных позвоночных начинается в заднем или среднем мозге, а оканчивается на эффекторных нейронах спинного мозга.
Нисходящие волокна переходят на противоположную сторону, как и восходящие волокна. Перекрёсты образуются как на уровне продолговатого мозга, так и в непосредственной близости от нейронов- мишеней спинного мозга (см. рис. II- 19, а-в). Спинному мозгу первичноводных позвоночных не свойственна широкая морфологическая изменчивость. Его строение сходно у различных видов, авыявленные отличия не могут служить причиной тупиковой эволюционной специализации. Это понятно из того, что спинной мозг является своеобразными рецепторно исполнительными воротами организма. Через них в мозг приходит информация о теле животного и выходят сигналы, управляющие внутренними органами и мускулатурой.
В связи с этим мозг организован предельно экономично и крайнеконсервативно. Достаточно упомянуть о том, что в процессе эмбриональной дифференцировки спинного мозга погибает до 85 % всех клеток, которые могут участвовать в работе этого отдела центральной нервной системы. Оставшиеся клетки успешно обеспечивают основные автономные и «воротные» функции, но ни о какой быстрой адаптивной изменчивости или морфологических перестройках речь не идёт.
Совершенно другое дело — строение головного мозга: 5 уже упомянутых отделов — передний, промежуточный, средний, задний с мозжечком и продолговатый мозг (см. рис. II-17, б) связаны с обслуживанием конкретных внешних и внутренних органов чувств.
Поскольку биология первичноводных животных крайне разнообразна, морфология этих отделов широко варьирует (см. рис. II-20; рис. II-21; II- 22). Изменчивость строения связана с адаптивными морфологическими перестройками основных центров головного мозга (Halpern, 1980; Foreman et аl., 1985).
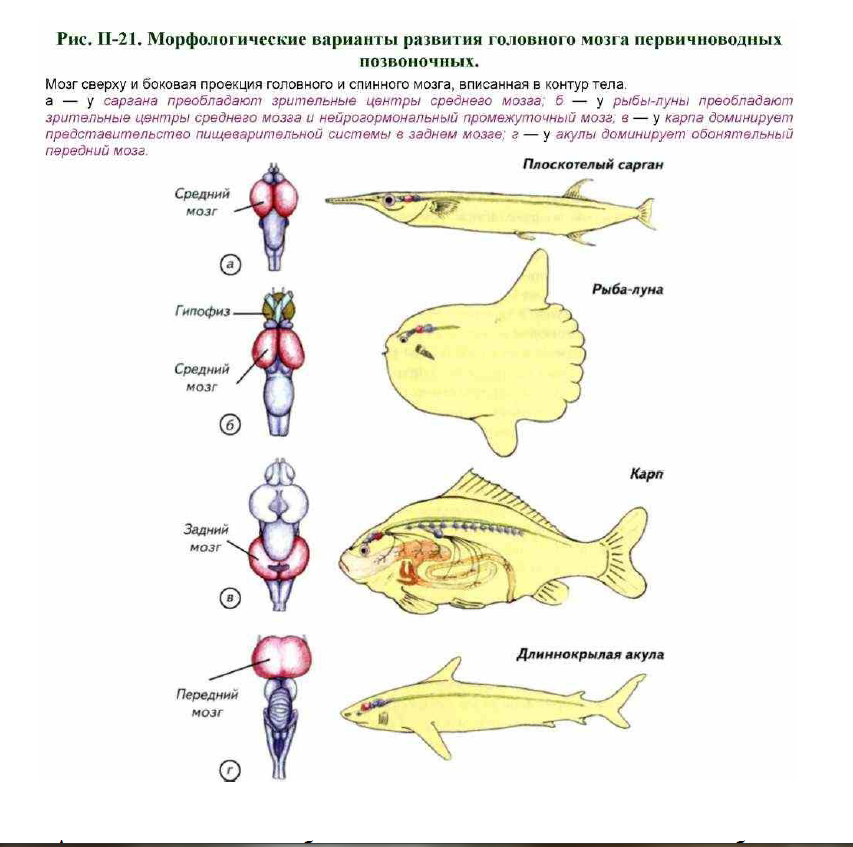
Передний мозг состоит из парных полушарий и является центром, обеспечивающим анализ химических сигналов из внешней среды (см. рис. II-17; II-18). Из органов обоняния поступают рецепторные сигналы, которые обрабатываются комплексом первичных и вторичных обонятельных центров. В переднем мозге происходит классификация сигналов, идентифицируется направление на источник запаха и формируется система обменных сигналов с другими отделами головного мозга. Понятно, что в водной среде запахи распространяются не так быстро, как в воздухе, но намного дольше сохраняются в окрестностях источника, что даёт определённые преимущества. Большинство первичноводных позвоночных имеет хорошо развитые органы обоняния.
Они позволяют определять расположение и движение пищевых объектов, направление миграции половых партнёров и конкурентов. В некоторых случаях органы обоняния достигают гигантских размеров. У многих пелагических акул передний мозг составляет примерно треть или даже половину переднего мозга (см. рис. II-18, б; II-21, г). Центры анализа обонятельных сигналов увеличиваются до таких размеров, что маскируют
разделение полушарий (Halpern, 1980).
Остальные отделы головного мозга относительно невелики, что позволяет рассматривать длиннокрылую акулу (Carcharhinus longimanus) как пример крайней специализации. У акул есть двойная ноздря. Одна ноздря служит для входа воды, а другая — для выхода. В зависимости от биологии акул орган обоняния промывается водой или при поступательном движении, как у длиннокрылой акулы, или при латеральных покачиваниях головой, как у колючей акулы.
Анатомические особенности строения органов обоняния предопределяют способности пелагических акул к обнаружению пищи на больших расстояниях. При поступательном движении у большинства серых акул (Carcharhinidae) вода захватывается входной ноздрей, как воздухозаборником самолёта. Это обеспечивает быструю смену воды и возможность захвата пузырьков воздуха. Воздушные пузырьки могут захватываться и удерживаться в органах обоняния, если при движении со скоростью больше 1,2 м/с рострум приподнимается на 1–2 см над поверхностью воды. Пузырьки воздуха удерживаются в органах обоняния при помощи специализированных перегородок в обонятельном мешке — ламелл и снижения скорости движения животного.
При задержке воздуха между ламеллами происходит растворение содержащихся в пузырьках веществ. Акулы не могут непосредственно рецептировать воздух и поэтому депонируют его в обонятельном мешке, дожидаясь повышения концентрации веществ вокруг пузырьков. Если вода не содержит
привлекательных запахов, то акулы набирают скорость и выдавливают водой пузырьки воздуха, расположенные между ламелл, содержащих рецепторные клетки. Затем цикл повторяется.
Надо отметить, что площадь поверхности рецепторных ламелл органов обоняния серых акул массой 75 кг может быть больше, чем у человека, в 60–90 раз. В воздухе над поверхностью океана запахи распространяются очень быстро, что позволяет акулам эффективно находить добычу.
Следовательно, длиннокрылая акула может анализировать химические сигналы как из водной, так и из воздушной среды. Для обслуживания столь развитой обонятельной системы у серых акул сформировался гипертрофированный передний мозг, а обоняние стало ведущей системой афферентации. Если экстраполировать на человека образ мира такой акулы, то он предстанет как богатая запахами, но почти тёмная комната, она будет разделена по вертикали на мир водных и мир воздушных запахов. Через запахи будет передаваться почти вся информация, хотя кожные механорецепторы, специализированные электрорецепторные органы и весьма слабое зрение будут дополнять «обонятельную» картину
мира (Halpern, 1980).
Следует отметить, что различия в строении переднего мозга проявляются как на количественном, так и на качественном уровне (см. рис. II-20, F). Если у пластиножаберных и американского чешуйчатника (Lepidosiren paradoxe) полушария переднего мозга замкнуты (см. рис. II-20, а), то у рыбы-луны передний мозг заметно эвертирован (см. рис. II-20, в).
Это означает, что полушария переднего мозга как бы вывернуты желудочками наружу. Некоторые костистые рыбы и рогозуб (Neoceratodus forsterl) имеют промежуточный вариант строения полушарий, как показано на рис. II-20, б. В эвертированных полушариях переднее сосудистое сплетение лежит на верхней поверхности мозга или частично заполняет полость черепа. При обычном строении полушарий сосудистое сплетение
располагается в полостях латеральных мозговых желудочков (Kappers, Huber, Grosby, 1936; Kardong, 1995).
Промежуточный мозг не связан с конкретными анализаторами, за исключением пинеального комплекса. Последний включает в себя нейроэндокринные центры и теменной фоторецептор (глаз), который воспринимает только уровень освещённости и ответствен за суточные ритмы организма.
В промежуточном мозге сосредоточены нейрогормональные центры, которые отвечают за рост, половое созревание, осмотический баланс, работу эндокринных органов, сезонную и суточную активность животного.
Через промежуточный мозг осуществляются связи между передним, средним, задним и продолговатым мозгом. У первичноводных животных уровень морфологического развития промежуточного мозга является своеобразным свидетельством определённой стратегии поведения животного. Если промежуточный мозг развит столь значительно, как у рыбы-луны, то механизмы регуляции поведения преимущественно гормональные (см. рис. II-21, б).
Действительно, трудно найти среди первичноводных позвоночных животное с таким гипофизом, который лежал бы ростральнее переднего мозга, а обонятельные и зрительные нервы проходили бы над ним. У рыбы- луны гипертрофия промежуточного мозга сочетается с незначительными размерами переднего мозга и преобладанием зрительной системы. Эти животные ведут подвижную жизнь в мировом океане, им свойственна гигантская плодовитость. За один раз рыба-луна может отложить до 300 млн икринок. Рыба-луна питается зоопланктоном и практически не пользуется обонянием. Зрение ей необходимо в период размножения, когда животные собираются в большие скопления.
Преобладание зрительных и гормональных центров над другими отделами мозга полностью соответствует биологии рыбы-луны. Таким образом, количественное доминирование нейроэндокринных центров промежуточного мозга над другими отделами центральной нервной системы является надёжным признаком гормонально-инстинктивного типа поведения животного (Foreman et al., 1985).
Зрительная система может стать ведущей системой афферентации у первичноводных позвоночных. В этом случае отмечается чрезвычайное развитие крыши среднего мозга — основной мозговой части зрительного анализатора. Примером такого строения мозга может быть средний мозг форели, чёрного марлина, летучей рыбы или плоскотелого саргана (см. рис. II-18, в, г, II-19, д; II-20, ж, и; II-21, а). Крыша среднего мозга образует крупные выпячивания, которые напоминают парные полушария переднего мозга, но это обманчивое впечатление исчезает, если рассмотреть III желудочек среднего мозга (см. рис. II-20, R). На сечениях видно, что реальных полушарий нет, а иллюзию создаёт симметричное латеральное расширение слоистой крыши (см. рис. II-19, д). Средний мозг у первичноводных животных является не только мозговым центром зрения.
В крыше среднего мозга сосредоточены представительства боковой линии, вестибулярного аппарата и органов электрорецепции, которые хорошо развиты у многих первичноводных позвоночных. В вентральной части среднего мозга лежат преимущественно двигательные центры черепно- мозговых нервов.
У большинства первичноводных позвоночных средний мозг выполняет функции принятия генерализованных решений. Основой для такой активности служит разнообразная сенсорная информация, которая поступает в средний мозг от внешних и внутренних анализаторов. У первичноводных позвоночных основой для принятия решения являются сенсомоторная и зрительная информация, сигналы от вестибулярной системы, органов боковой линии и обонятельной системы. Только последняя не представлена прямыми связями в крыше среднего мозга.
Практически все основные моторные ядра заднего и продолговатого мозга взаимодействуют с крышей среднего мозга у круглоротых, хрящевых и костистых рыб. У хрящевых и костных рыб они образуют прямые связи, идущие как от первичных моторных центров, так и от вторичных ядер, обслуживающих мозжечок. У круглоротых таких прямых связей намного меньше, чем у акул. Это связано с тем, что исторически более древним вариантом поступления сигналов в крышу среднего мозга является ретикулярный путь. По-видимому, первоначально сигналы проходили через ядра рострального края ретикулярной формации — покрышку среднего мозга, а только затем поступали в тектум.
Этот путь хорошо выражен у круглоротых и представлен небольшим количеством волокон практически у всех позвоночных. Однако у акул система сенсомоторно- тектальных связей значительно изменилась. Основные моторные центры установили прямые связи с тектумом, а тегментно-тектальные моторные пути стали вспомогательными.
У первичноводных позвоночных с функциональным преобладанием рения над другими дистантными анализаторами крыша среднего мозга стала центром принятия решений. Она стала выполнять функции, аналогичные роли ассоциативной коры переднего мозга млекопитающих.
Крышу среднего мозга можно назвать ассоциативным центром с большой натяжкой. Скорее это центр тотального сравнения различных раздражителей. Сравнительный анализ сигналов осуществляется следующим образом.
Каждая из сенсорных систем, за исключением обоняния, представлена условным анализаторным слоем в крыше среднего мозга. При этом соблюдается топическая эквивалентность представительства каждого анализатора. Это означает, что информация от органов боковой линии из средней части левой стороны тела рыбы приходит примерно в то же место, куда поступают сигналы от сетчатки левого глаза. В то же место, но в
другие слои крыши среднего мозга приходят сигналы от электрорецепторов и туловищной мускулатуры. Все сигналы сравниваются между собой специальными клетками, пронизывающими крышу среднего мозга по вертикали. Если один из сенсорных слоёв возбуждён больше, чем другие, то его активность становится ведущей.
Самая возбуждённая сенсорная система определяет выбор одной из инстинктивных программ поведения, но при этом постоянно происходит сравнительный контроль возбуждений от других органов чувств. Если источником максимального возбуждения становится другой слой, то он приобретает приоритет в подборе инстинктивного ответа на новую ситуацию.
Такая система принятия решения очень эффективна для небольших объёмов нервной ткани с относительно пропорциональным развитием мозгового представительства органов чувств, но ожидать сложного ассоциативного поведения от животных, «думающих» крышей среднего мозга, не приходится. Скорее это самая совершенная система для выбора оптимальной последовательности инстинктивных форм поведения. У
первичноводных позвоночных мозг недостаточно велик для глубокого анализа окружающего мира, а доля благоприобретённого индивидуального поведения редко превышает 5–7 %. Основой поведения является набор врождённых инстинктов, обеспечивающих как выживание, так и размножение. На нервную систему возлагается функция выбора программы поведения, наиболее адекватной для данной ситуации. Выбор осуществляется в крыше среднего мозга. Он происходит по принципу сравнения возбуждения от различных органов чувств.
Наиболее возбуждённая система имеет преимущество в выборе двигательного ответа на раздражение. Она запускает инстинктивную программу поведения, которая реализуется до тех пор, пока возбуждение другой рецепторной системы не «отнимет» право выбора. Смена центра, выбирающего новую поведенческую программу, происходит практически мгновенно, чем объясняется «немотивированная» смена активности почти всех первичноводных позвоночных. Следовательно, среднемозговой центр принятия решений построен по иерархическому принципу, но с динамической возможностью мгновенного возврата на исходную позицию и смены формы поведения. Такая конструкция мозга стала идеальной для длительной эволюции позвоночных. Максимального развития эта система выбора формы поведения достигла у рептилий. Их средний мозг
полностью реализовал ассоциативно-рефлекторные принципы, заложенные в эволюцию мозга первичноводных позвоночных.
Надо отметить, что у низших позвоночных крыша среднего мозга является местом принятия решения и, как следствие, источником генерализованного двигательного ответа. У первичноводных позвоночных и амфибий преобладают прямые нисходящие двигательные пути, которые оканчиваются в двигательных центрах заднего и продолговатого мозга.
При помощи этих связей осуществляется контроль над моторной активностью. В другие отделы мозга нисходящие волокна из среднего мозга направляются в крайне незначительном количестве.
Иным образом организованы сенсомоторные центры заднего и продолговатого мозга. В этих отделах мозга нет специального места для интеграции различных органов чувств. Все взаимодействия между сенсорными и моторными центрами осуществляются за счёт специальных отростков нервных клеток, которые образуют своеобразную ретикулярную (сетчатую) формацию. В заднем и продолговатом мозге в самом общем
виде сохраняется пространственная структура спинного мозга (см. рис. II- 19, а-г). В дорсолатеральной части этого отдела сосредоточены сенсорные центры, а в медиовентральной — моторные. Эти центры получают восходящие сигналы от спинного мозга и чувствительных черепно- мозговых нервов. Нисходящие двигательные волокна управляют большей частью скелетной мускулатуры и работой внутренних органов. У
большинства первичноводных позвоночных задний и продолговатый мозг образует специфическую полость, возникающую в результате эмбрионального разворачивания верхней стенки нервной трубки. Она носит название IV желудочка и соединена рострально с III желудочком, а каудально — с центральным каналом спинного мозга (см. рис. II-17; II-19).
Над дорсальной поверхностью IV желудочка формируется сосудистое сплетение, которое может достигать огромного размера и располагаться над дорсальной поверхностью всего мозга. Анатомическая организация дна и полости IV желудочка отражает адаптационные особенности первичноводных позвоночных. Примером могут служить представители семейства карпозубых. У обычного карася и карпа латеральные стенки заднего мозга чрезвычайно расширены (см. рис. II-20, м; II-21, в). Более того, в этих зонах сформированы стратифицированные структуры, которые напоминают крышу среднего мозга. Однако эти гигантские образования, зачастую доминирующие в головном мозге, представляют собой не что
иное, как разросшееся ядро блуждающего нерва (X). Иногда это разрастание называют вагальной долей заднего мозга. Действительно, его линейные размеры могут превышать некоторые отделы центральной нервной системы.
Понятно, что при таком преобладании в мозге представительства блуждающего нерва он и становится одним из основных центров при выборе той или иной программы поведения. Для карпа самой существенной информацией для выбора формы поведения является состояние его внутренних органов, в первую очередь пищеварительной системы. Карп оценивает эффективность действий по состоянию своего желудка, что становится решающей мотивацией в выборе конкретной поведенческой тактики.
В полости IV желудочка может быть увеличено представительство и других черепно-мозговых нервов. Примером могут служить многочисленные сомы, способные к дифференциальной соматической и вкусовой чувствительности при помощи специальных выростов на голове (усов). У них на дне IV желудочка морфологически может обособляться ядро лицевого нерва (VII). Подобное развитие соматической чувствительности приводит к столь значительному увеличению размеров этого центра, что он может визуально закрывать почти всю полость IV желудочка. К сходным последствиям приводит гипертрофированное развитие представительства языкоглоточного нерва (IX) (см. рис. II-20, л).
Такое интенсивное развитие ядер языкоглоточного нерва обычно для рыб с развитой системой генерации электрических разрядов. У электрического ската центры управления электрическими органами полностью закрывают полость IV желудочка.
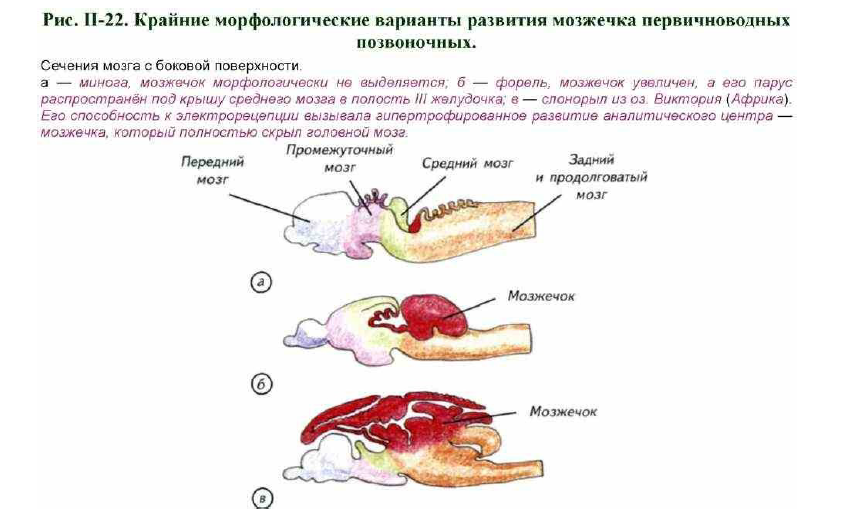
Наибольшим диапазоном изменчивости среди сенсомоторных центров мозга обладает мозжечок. Он расположен у переднего края заднего мозга и может достигать огромных размеров, закрывая собой весь головной мозг (см. рис. II-22). Его развитие зависит от нескольких причин. Наиболее очевидная связана с пелагическим образом жизни, хищничеством или способностью к эффективному плаванию в толще воды.
Наибольшего развития мозжечок достигает у пелагических акул (см. рис. II-18, б). В нём формируются настоящие борозды и извилины, которые отсутствуют у большинства костистых рыб (см. рис. II-18, в, г). В этом случае развитие мозжечка вызвано сложным движением акул в трёхмерной среде мирового океана. Требования к пространственной ориентации слишком велики, чтобы это не отразилось на нейроморфологическом обеспечении вестибулярного аппарата и сенсомоторной системы. Этот вывод подтверждается исследованием мозга акул, обитающих на билатерали и около дна. У акулы-няньки нет развитого мозжечка длиннокрылой акулы, а полость IV желудочка полностью открыта (см. рис. II-18, а). Её биология не предъявляет таких жёстких требований к пространственной ориентации, как у длиннокрылой акулы. Следствием стали относительно скромные размеры мозжечка. Для миксин и миног как пространственная ориентация, так и контроль за высокими скоростями движения не имеют важного
биологического значения (Dodd, Dodd, 1985). Являясь паразитическими животными или падальщиками, круглоротые не нуждаются в сложно координации движений, что отражает строение их мозжечка (см. рис. II- 22, а).
Однако размеры и форма мозжечка у первичноводных позвоночных могут изменяться не только в связи с пелагическим или относительно оседлым образом жизни. Поскольку мозжечок является центром анализа соматической чувствительности, он принимает самое активное участие в обработке электрорецепторных сигналов. Электрорецепцией пользуются очень многие первичноводные позвоночные. На сегодняшний день известно, что 70 видов рыб обладают развитыми электрорецепторами, а около 500 видов могут генерировать электрические разряды различной мощности. Примерно 20 видов способны как генерировать, так и рецептировать электрические поля. Наиболее изучена эта способность у гимнарха (Gymnarchus niloticus), рецептирующего электромагнитное поле, создаваемое им самим. При попадании в его поле объектов различной электропроводности гимнарх может определить направление их движения, размер и скорость.
Электрорецепция используется для ухаживания друг за другом особей различного пола и подавления электромагнитных полей конкурирующих особей или других видов. Кроме гимнарха, аналогичные способности генерировать и воспринимать собственные сигналы известны у других костистых рыб, акул и скатов.
Электромагнитные сигналы первичноводные позвоночные воспринимают при помощи рецепторов двух основных типов: ампульных (ампулы Лоренцини) и клубочковых. У некоторых видов присутствуют рецепторы обоих типов, но у большинства только одного. Ампульные электрорецепторы приспособлены для восприятия медленно изменяющихся электрических полей, а клубочковые реагируют на быстрые изменения, поэтому у активно плавающих рыб, обитающих в непрозрачной воде, более развиты клубочки, а у хищников в прозрачной воде — ампульные рецепторы. Если основной системой афферентации становится электрорецепция собственного электромагнитного поля или внешних электрических полей, то мозжечок начинает выполнять роль сенсорного мозгового центра. У всех рыб, обладающих электрорецепцией, мозжечок развит чрезвычайно хорошо (см. рис. II-22, в). Зачастую полушария мозжечка так велики, что закрывают с дорсальной поверхности весь мозг.
Таким образом, структурные отделы головного мозга первичноводных позвоночных представляют собой своеобразные маркёры морфофункциональной адаптации вида к определённым условиям обитания. Анализ организации нервной системы первичноводных позвоночных даёт объективную информацию о развитии систем афферентации, способах принятия решений и ведущих мотивационных центрах головного мозга. Однако не менее интересен эволюционный путь возникновения столь компактной и эффективной системы управления поведением. Палеонтологических свидетельств возникновения
современной конструкции мозга первичноводных позвоночных крайне мало. Даже самые древние находки содержат в основном информацию об уже сложившемся современном типе организации головного и спинного мозга. Реконструируя становление нервной системы первичноводных позвоночных, приходится опираться на архаические черты строения мозга современных видов.
По-видимому, появление древних хордовых не сразу привело к заметным изменениям в биологии водной среды. Судя по всему, первые хордовые были относительно небольшими животными, размером от нескольких сантиметров до полуметра. Они явно не могли составить серьёзной конкуренции процветавшим водным беспозвоночным, которые зачастую были намного больше, чем молодая группа хордовых. По размерам древние позвоночные явно проигрывали беспозвоночным и не могли на равных конкурировать с ними. Нервная система не давала особых преимуществ этой новой группе, скорее наоборот. Небольшая и хорошо детерминированная нервная система с набором эффективных поведенческих программ давала беспозвоночным заметные преимущества в конкуренции с древними хордовыми. Нервная система древних позвоночных обладала только одним положительным качеством — способностью к почти неограниченному увеличению своих размеров.
Однако это преимущество было реализовано далеко не сразу. На первом этапе хордовые решали проблему конкуренции с беспозвоночными при помощи выбора среды обитания, становления строения мозга и скелета.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы