2. Тропизмы и таксисы простейших
Относительно небольшие размеры простейших дают возможность непосредственно использовать рецепторный аппарат мембраны для быстрого изменения поведения. Размер большинства простейших обычно не превышает нескольких миллиметров (рис. I-2). Это не значит, что нет более крупных одноклеточных. Известны виды, достигавшие нескольких десятков сантиметров, но поведенчески они были столь же пассивны, как и растения. Однако небольшие одноклеточные дают наиболее выраженный пример сложного и даже рефлекторного поведения. Сенсорный аппарат простейших сходен с таковым растений, но цитоплазматически намного более специализирован. Общая химическая чувствительность простейших хорошо известна. Простейшие прекрасно двигаются по градиенту пищевых растворов и избегают растворов щелочей, кислот и солей. Присутствие в растворах катионов калия, лития, натрия и аммония вызывает реверсию активности ресничек и жгутиков, которые служат для передвижения многих простейших. Надо отметить, что положительный и отрицательный хемотаксис инфузорий лёг в основу теории поведенческих тропизмов Ж. Леба.Тропизмами, или таксисами, Ж. Леб называл простые реакции растений и животных на свет, химические вещества, источники электромагнитных полей и т. д. Например, если животное двигалось в сторону света, Ж. Леб называл это положительным фототропизмом, если от света — отрицательным. Наиболее популярными были исследования, связанные с гелиотропизмом (движение к солнцу или от него), термотропизмом (движение к теплу или от него), хемотропизмом (движение к веществу или от него), геотропизмом (чувствительность к направлению гравитационных сил), тигмотропизмом (чувствительность к механическому контакту), цитотропизмом (стремление клеток к адгезивному слипанию или разделению) и реотропизмом (движение в потоке воды). В основе учения Ж. Леба лежит универсальная способность цитоплазмы клеток всех живых организмов к раздражению. Поскольку этим свойством обладают все клетки, Ж. Леб делает вывод о единстве животных и растений, а заодно и о незначительной роли нервной системы. Он совершенно серьезно писал: «У животных, обладающих нервами, явления гелиотропизма вызываются теми же причинами (формой тела и раздражимостью протоплазмы клетки), как и у растений, не имеющих нервной системы. Таким образом, явления тропизма не могут основываться на специальных свойствах центральной нервной системы…».
Несмотря на всю оригинальность таких представлений, теория Ж. Леба нашла многочисленных сторонников. Его последователи начали проверять раков на «хининовый таксис», червей на «солевой таксис», лягушек и рыб на «электрический таксис». Возникла даже разветвлённая система понятий, которые предусматривали различение таксиса и тропизма. Тропизмом стали называть изменение ориентации или направления роста прикреплённого организма. Обладателями тропизмов считали растения. Под таксисом подразумевали уже самодвижущийся организм, который или удалялся, или приближался к источнику раздражения (Iftode, Prajer, Frankel, 2001; Barbanera, Erra, Banchetti, 2002). Понятно, что такие «жизненно важные» таксисы и тропизмы не только ничего не добавляли к психологии, но и превращали зоопсихологию в теолого-лингвистическую науку. Сторонники теории тропизмов не остановились на уравнивании тропизмов животных и растений. Они успешно доказывали, что зрительное восприятие человека ничем не отличается от фототропизма инфузорий. Смысл этих доказательств состоял в поиске универсальной единицы поведения — «атома», который они называли тропизмом, или таксисом. Ж. Леб и его последователи считали, что, найдя некоторое количество универсальных единиц поведения, они смогут «вычислить» или «расчленить» любое сложное поведение животного и мышление человека.
Несмотря на все заблуждения и фантазии, сторонники таксической теории Ж. Леба были хорошими экспериментаторами. Работая на одноклеточных организмах, они привлекли огромное внимание к изучению их биологии и поведения. В многократно повторённых экспериментах было установлено такое интереснейшее свойство одноклеточных организмов, как привыкание. Эти эксперименты проводили на парамециях, которых предварительно приучали к определённой температуре, а затем помещали в ванночку с температурным градиентом. Оказалось, что оптимальной для парамеций является температура 24–28 °C.
Рис. 1-2. Микроскопические пресноводные организмы.

На уровне микроскопических размеров преимущества нервной системы почти незаметны, что позволяет одноклеточным успешно конкурировать с многоклеточными организмами, обладающими примитивной нервной системой, а-в — пресноводные гидры с диффузной нервной системой, а — гидра; б — гидра после прикосновения к ней; в — гидра в спокойном состоянии.
Однако при выдерживании инфузорий при низких температурах они предпочитали уменьшение обычного оптимума. Экспериментаторы рассматривали такие результаты как явное свидетельство «обучаемости» парамеций.
Универсальность тропизмов и таксисов как единиц поведения подчёркивалась в популярных опытах с использованием электрических полей и токов. Гальванотаксисы и тропизмы изучали почти на всех живых существах. Личинки амфибий и молодь костистых рыб ориентируются вдоль направления тока и головой к аноду. Моллюски, членистоногие и черви прекрасно чувствуют электрические поля и ориентируются в них к одному из полюсов. Обезглавленные черви стараются избегать «напряжённого» участка земли. Эксперименты с одноклеточными организмами показали, что они избирательны по отношению к электромагнитному полю или токам в водных растворах. Парамеции и амёбы предпочитают отрицательный полюс, а другие инфузории — положительный. Некоторые одноклеточные организмы не проявляют гальванотаксиса, но ориентируются вдоль или поперёк электромагнитного поля. Опыты на растениях показали существование аналогичного гальванотропизма. Корни, развивавшиеся в условиях электронапряжённости почвы, поворачивались в сторону отрицательного полюса, проявляли положительный гальванотропизм. Таким образом, универсальные рецепторные свойства клеток позволяют увидеть нейроподобные эффекты как у животных, так и у растений.
Однако реакции простейших построены далеко не на общей клеточной чувствительности, характерной для растений. У простейших известны и сложные специализированные рецепторы — органоиды. Такими рецепторами являются ложноножки корненожек Allogromia sp. или осязательные волоски трубача Stentor sp. В качестве особой фоторецепторной системы формируются глазки, или стигмы. Эти образования пигментированы, позволяют простейшим ориентироваться в градиенте света и дифференцировать различные длины волн светового диапазона. Описаны эксперименты, в которых удавалось выработать рефлекторное поведение парамеций на красный и синий свет. Стигмы особенно хорошо развиты у динофлагеллат, эвглен и фитофлагеллат. Следует отметить, что среди простейших существуют группы (Amoeba), прекрасно реагирующие на свет, но не имеющие специализированных мембранных органелл для его восприятия (Meglitsch, Schram, 1991). Особо развиты у простейших органоиды движения. Небольшой размер организма позволяет эффективно пользоваться приспособлениями ультраструктурного размера для перемещения, питания или агрессии. Корненожки и амёбы образуют мембранные образования, называемые ложноножками, или псевдоподиями. Они позволяют многим простейшим двигаться и захватывать пищу. Псевдоподии крайне разнообразны. Выделяют широкие, цилиндрические и округлые на конце — лобоподии, длинные и нитевидные — филоподии, сетчатые, разветвляющиеся и анастомозирующие — ретикуло-подии. Существуют и более эффективные в движении псевдоподии со стержнем внутри — аксоподии. Они способны не только вытягиваться и втягиваться, но и сокращаться (McConnel, 1966). Для движения простейшие используют довольно крупные жгутики или многочисленные мелкие реснички, которые позволяют перемещаться с невероятной скоростью. У простейших существуют аналоги мышечных волокон — мионемы, которые позволяют изменять форму тела или совершать сложные движения. Сжатие стебелька у сувойки представляет собой результат мышечного сокращения.
Следовательно, простейшие организмы способны к автономному питанию, размножению, движению и адаптации к изменяющимся условиям внешней среды. Наличие эффективных органов движения является принципиальным отличием простейших от растений. Если есть активное движение, значит, можно наблюдать и быстрые поведенческие реакции. Их существование свидетельствует о механизмах системного управления простейших, которые построены по аналогии с нервной системой, тем более что при отсутствии нервной системы они проявляют прекрасную обучаемость.
С момента признания существования простейших в 40-х годах XIX в. началось изучение их поведения. Открытие мира простейших привело к известному буму в наблюдениях, а любители «наливок» сформировали целое направление. «Наливочниками» называли исследователей, разводивших микроорганизмы в настоях или наливках различного состава. Название возникло от латинского infusum — отвар, настой. Доступность и разнообразие объектов завораживали первых микроскопистов, что привело к появлению многочисленных описаний поведения простейших. Одним из первых наблюдателей мира простейших был Р. фон Розенгоф. Ещё в 1755 г. он детально описал свободноживущую амёбу и патетически назвал её микроскопическим Протеем.
Сложность поведения простейших поразила первых исследователей и привела к переоценке их психических способностей. Так, В. Кюне в 1859 г. считал колокол сувойки головой. При отрезании этой «головы» стебелёк сувойки переставал сокращаться, что позволило заподозрить у этих животных сложную психику. В то время считалось, что сувойки способны к ощущениям, распознаванию, обладают сознанием и совершают волевые поступки. Работы тех лет полны интересными свидетельствами личной жизни одноклеточных организмов. Психические переживания простейших описывались как «приятные ощущения», а питание амёб происходило «с чувством удовольствия». Авторы без тени сомнения наделяли простейших собственными гастрономическими переживаниями и ощущениями. Понятно, что эти смелые фантазии возникли на пике увлечения изучением поведения простейших и связаны с быстрыми реакциями на различные физико-химические воздействия.
Эти примеры лишний раз подчёркивают, что у любого живого организма можно увидеть признаки сложного поведения. Для этого достаточно рецептивного аппарата растений в сочетании с быстрым ответом любых эффекторных систем. Все аналогичные события происходят и у растений, но скорость процессов так отличается от темпов нашей жизни, что быстрые, с позиций растений, ответы на внешние воздействия мы не замечаем.
Тем не менее сложность адаптивных реакций одноклеточных свободноживущих организмов заслуживает дополнительного внимания. Проведено несколько блестящих экспериментов, которые демонстрируют реальную способность простейших к поведенческой адаптивности и создают видимость способности к примитивному обучению.
Свободноживущие амёбы проявляют весьма разнообразные стратегии захвата пищи, реагируют на свет и колебания воды. Если рассмотреть при помощи микроскопа поведение амёбы, то возникает уверенность в отнюдь не физико-химической природе питания простейших. Амёба, столкнувшись с большим или незакреплённым пищевым фрагментом, демонстрирует разнообразные приёмы его захвата. Сначала она использует одиночную псевдоподию для охватывания пищевого фрагмента. Если фрагмент ускользает, то начинается его охватывание с различных сторон двумя, а иногда и тремя псевдоподия-ми. Каждая неудача в захвате пищи вызывает применение нового приёма, разнообразие которых весьма велико. Этот пример показывает, что амёбы корректируют своё поведение в зависимости от результата.
Прикреплённые инфузории ещё более удобны для наблюдений и элементарных поведенческих экспериментов. Широко известны и многократно проверены результаты опытов с трубачом (Stentor roeseli), которого посыпали растёртым кармином. Обычно выделяют 4 этапа в реакциях трубача на попадание кармина в воронку. Мне удалось наблюдать три из них. Первоначальная реакция на частицы кармина отсутствует, а затем трубач смещает воронку в сторону от падающих частиц. Если это не помогает, то он начинает активно работать околоротовыми ресничками, пытаясь выбросить уже попавшие в воронку частицы и предупредить попадание новых. При продолжении воздействия трубачи интенсивно сокращают свой стебелёк и в конце концов отрываются от поверхности и уплывают в другое место. Важна не сама цепь разнообразных адаптивных реакций, а обнаруженная «память» трубачей при прерывании воздействия. Если трубач не уплыл, а воздействие прекращено, то животное ненадолго запоминает ситуацию. При возобновлении посыпания кармином трубач уже не демонстрирует все стадии рецептивной адаптации. Он сразу начинает с прерванной стадии и обычно уплывает. Следовательно, инфузории обладают формой донервной памяти, которая сохраняется несколько десятков минут.
В начале XX в. в США были придуманы изящнейшие эксперименты по индивидуальному обучению инфузорий (Paramecium). Эти результаты впечатлили Херрика (C.J. Herrick), который убедился в их достоверности и привёл в своей монографии, изданной в 1924 г. Суть этих опытов довольно проста. Инфузорию-туфельку помещают в узкую стеклянную трубочку, где ей трудно развернуться. Инфузория доходит до конца закрытой трубочки и останавливается. Двинуться в обратном направлении она может только развернувшись. Инфузория начинает разворачиваться, предпринимая множество неудачных попыток. До первой удачи она затрачивает в среднем от 10 до 25 попыток. После 4–6 поворотов инфузория обучается разворачиваться с первой попытки.
Казалось бы, из этих экспериментов следуют два вывода: инфузории могут обучаться и запоминать выученные движения. Спустя почти 60 лет после первых опытов с поворотом и выходом из трубки началась новая серия опытов по обучению инфузорий (Huber, Rucker, McDiarmid, 1974). Четыре независимые группы экспериментаторов убедились, что тренировка парамеций является артефактом, а не реальным научением. В качестве демонстрации памяти часто приводится способность многих простейших собирать иглы губок для защиты своего тела. Поскольку дочерние организмы являются частью тела родителей, вместе с унаследованной цитоплазмой и ядром они усваивают и способность собирать иглы. Так аргументируется способность простейших к использованию памяти родителей.
Известны опыты с инфузориями по выработке кратковременных рефлексов на свет различной длины волны и интенсивности, постоянные электромагнитные поля, температурные градиенты и электрическое раздражение. Во всех случаях удавалось добиться кратковременного запоминания ситуации и её воспроизведения в схожих условиях. Это привело к спорным выводам о существовании у простейших условных ассоциаций. Наиболее спорные опыты заключались в обучении парамеций воспринимать сочетанные раздражители (McConnell, 1966). В сосуд с парамециями опускали серебряную иглу, покрытую бактериями. Парамеции собирались вокруг неё и поедали бактерий. Понятно, что вместе с бактериями в воде появлялись и ионы серебра. Парамеций кормили таким способом несколько раз, а затем опускали в сосуд иглу без бактерий. Парамеции всё равно собирались вокруг иглы. Они воспринимали присутствие ионов серебра в растворе как сигнал к кормёжке. Эта поведенческая реакция постепенно угасала, если иглу не покрывали бактериями. Однако такое поведение спонтанно восстанавливалось спустя продолжительное время. Повторные опыты и анализ методов проведения экспериментов показали, что желаемое выдаётся за действительное, а парамеции не могут научиться воспринимать сочетанный сигнал. Тем не менее Д. Дьюсбери в своей монографии (1978, перевод 1981) умудряется обсудить неудачные эксперименты в подглавке, посвященной ассоциативному научению простейших. Р. Шовен (1972), описывая упомянутые опыты, без тени сомнения признаёт у простейших способность к обучению и восприятию условных стимулов. Многие зоопсихологи в явной или скрытой форме допускают существование у простейших некоего механизма научения, ассоциативного поведения, памяти и т. д. Это напоминает неудержимые фантазии Э. Геккеля, который ещё в 1876 г. убеждал зоологов в существовании «души» в клетках. Требуя рассматривать простейших как объекты психологии, он писал: «Клетка- душа в монистическом смысле есть общая сумма энергий, заключённых в протоплазме; она нераздельна от клеточного вещества, как душа человека от нервной системы».
Во времена Э. Геккеля было достаточно здравомыслящих исследователей поведения простейших, которые использовали незатейливые приёмы для выяснения природы адаптивных реакций одноклеточных. Логика исследования была такова: строение простейших не позволяет индивидууму сделать понятие о своём объединённом целом, а отсутствие самосознания исключает возможность высших психических процессов. В связи с этим популярным опытом было разрезание простейших на небольшие части. Разрезанная на части «индивидуальность» простейших даже без ядер продолжала проявлять ту же самую активность, что и целый организм. Эти результаты позволили ещё в конце XIX в. Д.А. Томсону (1892) справедливо писать: «… Психические процессы не существуют у протистов, или же они неразрывны с молекулярными изменениями, которые совершаются в частях физического вещества». В настоящее время получены сотни мутаций Paramecium aurelia, которые были исследованы при помощи поведенческого, электрофизиологического и генетического анализов. В этих работах показаны физико-химические механизмы адаптивного поведения простейших, а не скрытый интеллект этих организмов (Iftode, Prajer, Franke! 2001).
Не удалось установить индивидуальное научение и при размножении простейших. Наиболее сложный половой процесс известен у инфузорий. Они могут размножаться без полового процесса, но иногда осуществляется конъюгация, приводящая к обмену генетическим материалом между особями. Это происходит нечасто, так как в обычной ситуации деление инфузорий осуществляется без конъюгации, а дочерние организмы являются полноценными клонами — копиями исходной особи. Интересно, что существуют не до конца исследованные «поведенческие» механизмы предотвращения инцеста — конъюгации (обмена генетическим материалом) с близкородственными особями. Ещё в 1885 г. одному исследователю удалось изолировать одиночную инфузорию (Stylonychia) и непрерывно проследить её поведение и размножение на протяжении 5 мес. За это время прошло 250 поколений инфузорий без единой конъюгации. Это привело к тому, что инфузории измельчали, ядрышко подверглось изменению, и они стали вымирать. При их объединении с особями из других клонов нормальный фенотип и поведение восстанавливались в первом же поколении.
Объединив результаты различных наблюдений и опытов, можно суммировать набор донервных способов контроля поведения простейших. Они могут рецептировать все основные типы внешних воздействий и отвечать на полученные сигналы различной активностью: пищевым поведением, размножением или движением по градиенту сигнала. Благодаря длительности биохимических реакций они могут создавать иллюзию «запоминания» индивидуального опыта. Это поведение проявляется в привыкании к определённым типам воздействий или избегании раздражителя. После завершения инерционных биохимических процессов псевдопамять исчезает без какого-либо следа. Следовательно, инерционная биохимическая память одноклеточных является своеобразным предшественником нейронной памяти многоклеточных.
Используя этот набор этологических и экспериментальных данных, можно сформулировать общую модель развития донервного поведения. Для наглядности её интересно представить графически (рис. I-3). Растения и животные, не имеющие нервной системы, обладают развитой чувствительностью ко всем основным типам сигналов: химическим, механическим и электромагнитным. Эти воздействия вызывают реакции довольно быстро: в течение долей секунды у простейших и секунд/минут у растений. При отсутствии нервной системы возможен выбор основного раздражителя. Скорость наблюдаемой реакции при выборе основного направления ответа зависит от размера организма. Простейшие реагируют быстро и очевидно, а растения медленно, что маскирует их высокую чувствительность к внешним влияниям. Краткосрочная память имеется у всех организмов, но она построена на биохимических внутриклеточных процессах.
Рис. I-3. Донервная интеграция ответов на внешние воздействия у растений (точки) и свободноживущих простейших (пунктир).
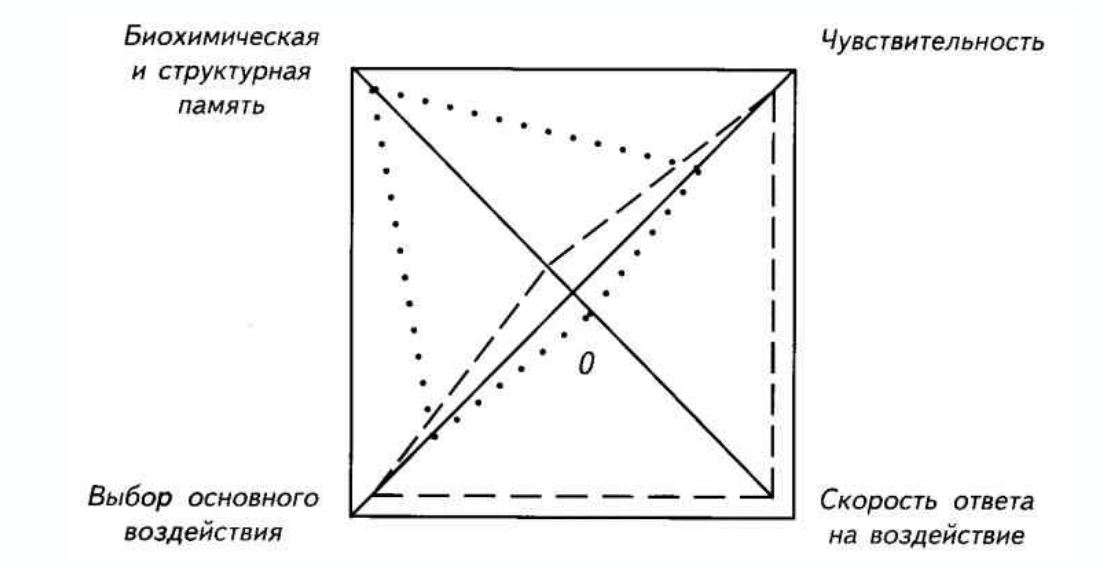
Различия сводятся к скорости ответа на воздействие, которая зависит от размера тела. Скорость реакций высокая у простейших, но у растений есть структурная «память», сохраняющая результаты предыдущих воздействий. Площади образовавшихся полей могут рассматриваться как критерии оценки направлений адаптивности донервных организмов.
Это делает невозможным использование механизмов индивидуального научения. Следовательно, основным принципом донервного взаимодействия с внешней средой является общая клеточная чувствительность, которая имеет избирательный характер. Общая клеточная чувствительность построена на трёх различных рецепторных механизмах: механочувствительности, хемочувствительности и электромагнитной рецепции. Однако эти условные «сенсорные варианты восприятия мира» далеко не равноценны. Надо подчеркнуть, что иллюзию осознанности поведения одноклеточных формирует скорость ответа на неспецифические влияния. Основные различия построены на скорости реакции при внешнем воздействии на организм. Если организм по сравнению с нами маленький и реагирует быстро, то нам кажется, что реакции осознанны и сходны с поведением животных, имеющих нервную систему. Отсутствие оперативной индивидуальной памяти компенсируется генетически детерминированными реакциями или направленным морфогенезом. Если бы растения и простейшие располагали индивидуальной памятью, они были бы самыми опасными существами на планете. К нашему счастью, для этого нужна нервная система.
Зачем нужна нервная система? Нервная система нужна не всем. Она не нужна тем, кто будет тысячелетиями неподвижен. Их «быстрый ответ» на внешние воздействия растягивается на десятки лет, им не надо ни быстрой реакции, ни мгновенной перестройки организма. Растения решили свои проблемы при помощи автотрофности, размера и времени жизни. Существует и другой вариант жизни без нервной системы. Можно поселиться в чудесном месте, где много пищи, организм защищен и согрет. Жизнь солитёра вполне соответствует этим требованиям. Солитёр и растения не обладают нервной системой. У растений нервной системы никогда не было, а у солитёра она полностью редуцирована. В обоих случаях её функции выполняют отдельные клетки, обладающие химической, электромагнитной и механической чувствительностью. Действительно, в оптимальных условиях питания и размножения нервная система вообще не нужна. Однако завидная для многих приматов судьба паразитических червей скорее исключение, чем правило в животном мире. Для большинства организмов окружающий мир слишком нестабилен и требует постоянного приспособления к нему. Адаптация к изменяющимся условиям должна быть генерализованной и охватывать все структуры организма. Таким органом быстрого и интегрированного реагирования стала нервная система.
Нервная система нужна тем, кто быстро двигается, активно вступает в контакт с разнообразными условиями внешнего мира и вынужден постоянно приспосабливаться. Быстротечный конформизм — причина формирования нервной системы. Действительно, нервная система малоподвижных организмов окажется невероятно упрощённой по сравнению с нервной системой активных животных. Актинии, асцидии, малоподвижные моллюски с крупными раковинами, коралловые полипы и многие другие животные имеют несложную нервную систему (рис. 1–4, а). У животных, прикреплённых к конкретному месту и занимающихся фильтрацией или захватом проплывающей пищи, очень простые задачи. Первая — рецепция пищевого объекта, вторая — его захват и переваривание. Для этого достаточно простых контактных рецепторов и органов удержания пищи, что, собственно, мы и видим у свободноживущих гидр и полипов. Их диффузная нервная система имеет небольшое окологлоточное нервное кольцо, которое и интегрирует несложные рефлексы.
Тем не менее эти простые реакции протекают на несколько порядков быстрее, чем морфогенетические перестройки тела у растений равного размера. Иначе говоря, нервная система необходима тогда, когда большому организму надо быстро адаптироваться к изменяющейся ситуации. Неважно, какая это ситуация: передвижение, захват пищи или избегание опасности, главное — скорость ответа на полученное раздражение (информацию).
Для осуществления быстрой интеграции работы крупного много- клеточного организма нужна не просто неспецифическая чувствительность живой клетки. Остро необходим и механизм передачи сигнала на большие расстояния. Нервные клетки обладают такими свойствами.
Рис. I-4. Два минимальных варианта организации нервной системы из функциональных блоков для однотипных, но дифференцированных ответов на различные типы внешних воздействии.
а — идеализированная схема — подразумевает разделение только сенсорных и двигательных функций, которые могут осуществляться даже одной сенсомоторной клеткой. Одни её отростки могут рецептировать раздражение, другие — передавать сигнал к эффекторным органам. Не исключено, что такие клетки обладали способностью к самостоятельному сокращению и рецепции.
б — более сложный набор неврологических блоков. В него входят сенсорный блок, блок сравнения сенсорных сигналов и эффекторный блок. Эти функции могут быть разделены между морфологическими структурами нервной системы или выполняться общей слабодифференцированной нервной сетью.
Они могут не только реагировать на различные воздействия, но и генерировать электрические сигналы. Импульсы формируются в аксонных холмиках и передаются по аксонам нейронов на значительные расстояния (см. рис. 1–4, б). Скорость проведения такого импульса может составлять от 0,13 м/с у актиний до 120 м/с в А-волокнах у человека. Генерация электрических сигналов и их распространение по поверхности мембраны нейронов — фундаментальное свойство нервной ткани животных. Однако при незначительном размере нейронов необходимо передавать этот сигнал от одной клетки к другой. Это было обеспечено увеличением размеров нервных клеток и их отростков. Нервные клетки имеют размер от нескольких микрон до нескольких метров. Самые большие клетки обнаружены в моторных отделах спинного мозга китообразных. Нейроны — самые крупные клетки животных, передающие информацию со скоростью около 400 000 км/ч.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы