24. Эволюция ганглиозной нервной системы
На заре эволюции многоклеточных сформировалась группа кишечнополостных с диффузной нервной системой (см. рис. II-4, а; рис. II- 11, а). Возможный вариант возникновения такой организации описан в начале этой главы. В случае кишечнополостных отдельные нейроны объединены в элементарную нервную сеть, которая относительно равномерно распределена между внутренним и наружным слоем клеток.
При всей простоте эта конструкция оказалась достаточно эффективной, и её обладатели дожили до настоящего времени. Обычная для Европы пресноводная гидра является классическим образцом поведения животного с такой нервной системой (см. рис. I-2). Из этой стартовой позиции начался гигантский эволюционный эксперимент по созданию эффективной системы управления организмом.
На первом этапе эволюции произошёл наиболее простой вариант усложнения нервной системы — концентрация нервных клеток в ганглии.
Процесс объединения нервных клеток шёл различными путями: во- первых, вокруг глотки, что характерно для всех без исключения животных того периода, во-вторых, вдоль несимметричного тела.
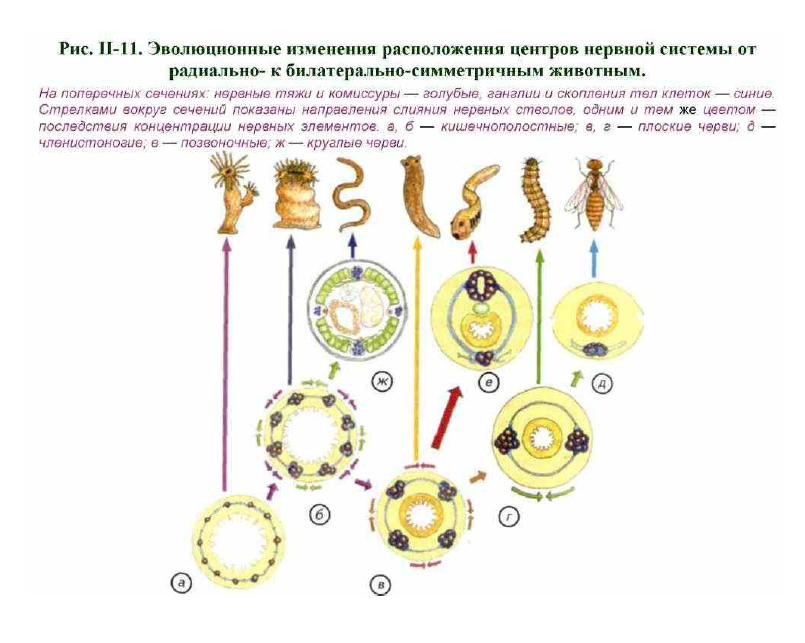
По выделенным тяжам нервных клеток проще проводить общий для всего тела управляющий сигнал. Это произошло уже у кишечнополостных, и такие животные дошли до нашего времени. У многих современных актиний есть «скоростные» морфологически выделенные нервные стволы (см. рис. II-4, в). Появление таких стволов с признаками параллельных ганглиев произошло в результате объединения соседних нейронов. Для
огромного большинства кишечнополостных такого усложнения диффузной нервной системы оказалось более чем достаточно. Они достигли того неврологического максимума, который при их образе жизни гарантировал питание и размножение. Однако даже среди радиально-симметричных кишечнополостных этот процесс происходил неодинаково. По-видимому, возникали как продольные, так и поперечные стволы из нервных клеток (см. рис. II-4, г, д).
Судя по всему, достижением стало появление радиально- комиссуральной конструкции, которая могла сочетать в себе как дифференциальные сегментарные реакции, так и генерализованные ответы на раздражения (см. рис. II-5, в; II-11, б). Радиально-комиссуральная нервная система могла стать основой для появления любых сегментированных существ. Это связано с тем, что отдельные сегменты имели собственные комиссуры, что позволяло решать сегментарные «проблемы» на месте, за счёт автономных нервных механизмов. Организм из повторяющихся сегментов становится менее уязвимым. Способность к масштабной регенерации — не менее важное свойство, чем эффективность питания или размножения. Из такого организма могли произойти самые разнообразные билатерально-симметричные конструкции (см. рис. 11, в, ж).
Причиной этих процессов могли быть дальнейшая концентрация нервной системы в виде стволов и комиссур. В одном случае стволы могли сливаться дорсально, латерально и вентрально, как показано на рис. II-11, б зелёными стрелками. Результатом стало появление 4 продольных нервных стволов (см. рис. II-11, ж). Один ствол лежит дорсально, два — латерально и один — вентрально. Возникла своеобразная билатеральная симметрия, которая относительно редко встречается у других беспозвоночных. Такая нервная система характерна для нематод. По-видимому, комиссуральные тяжи у нематод исчезли уже вторично, в связи с упрощением их биологии (см. рис. II-5, б). Аналогичное, но альтернативное слияние нервных стволов показано фиолетовыми стрелками (см. рис. II-11). Оно могло произойти у ползающего в придонном слое организма. После слияния нервных стволов возникла конструкция, похожая на нервную систему нематод, но нервные стволы расположены иначе: 4 нервных ствола лежат попарно, 2 сверху и 2 снизу. Нервная система такого строения встречается у многих плоских червей и хитонов (см. рис. II-5, г, II-10, б; II-11, в). Дальнейшая концентрация нервных элементов могла пойти по пути, отмеченному коричневыми или красными стрелками. При слиянии нервных стволов по принципу, отмеченному коричневыми стрелками (см. рис. II-11, в), возник вариант строения, характерный для большинства современных плоских червей. Это уже лестничная нервная система с симметричными парными нервными тяжами, идущими вдоль всего тела (см. рис. II-11, г). Её широкая распространённость среди беспозвоночных говорит сама за себя. Однако самое существенное состоит в том, что этот тип организации дал начало нервной системе членистоногих и сходных с ними животных (см. рис. II-д).
Слияние двух продольных нервных стволов произошло в вентральной части тела, как показано на рис. 11, г зелёными стрелками. Следы незавершенности этого процесса встречаются у многих современных членистоногих (см. рис. II-6; II-7). Возникновение брюшной нервной цепочки и специализированных головных ганглиев стало решающим событием в развитии животного мира. С такой нервной системой, конечно, невозможно творческое мышление в нашем понимании. Однако животным с небольшой продолжительностью жизни, развитой системой инстинктивных форм поведения, эффективной стратегией размножения и скромными линейными размерами в развитом интеллекте нет особой нужды. Проблемы выживания решаются не менее эффективно другими путями. Баланс между потенциальными возможностями инстинктивного поведения, ограниченной памятью и совершенством органов чувств был достигнут в ограниченном объёме головных ганглиев и скромной брюшной нервной цепочке. Это привело к
распространению членистоногих и сходных с ними групп в мировом океане, освоению пресноводных водоёмов, почвы и суши. Плодами процветания стали поведенческое разнообразие, самое большое число видов и сложнейшая биология представителей этой группы. Не менее интересен другой вариант слияния нервных стволов, показанный на рис. II-11, в красными стрелками. В этом случае произошло
объединение парных дорсальных и парных вентральных нервных тяжей. Результатом такой концентрации нервных стволов могло стать весьма оригинальное беспозвоночное с билатеральной симметрией и двумя нервными стволами: дорсальным и вентральным. На это могут претендовать отдельные щупальца иглокожих (Echinodermata).
Однако строение их нервной системы имеет мало общего с другими рассматриваемыми конструкциями. Иглокожие представляют собой интереснейшую в неврологическом отношении группу. Основная часть их нервной системы — околопищеводное нервное кольцо, поскольку о голове говорить не приходится. От него тянутся нервные стволы вдоль амбулакральной системы (по нижней части щупальца). Выраженных ганглиев нет, а нейроны расположены в стволах диффузно. Этот отдел нервной системы иглокожих носит название эктоневрального. Кроме него, у голотурий, офиур, морских звёзд и ежей встречается гипоневральная нервная сеть, а у морских лилий — ещё и апикальная. Все три параллельные нервные сети одновременно хорошо развиты только у офиур. Специализация офиур столь высока, что гипоневральная сеть имеет даже небольшие ганглиозные утолщения по ходу нервных стволов. В остальных группах обычно развита гипоневральная нервная сеть, которая часто анатомически сливается с эктоневральной сетью, как у голотурий.
Считают, что апикальная и гипоневральная сети выполняют моторные функции, а эктоневральная — сенсорные. Это не совсем так, поскольку разбросанные по телу осязательные клетки и светочувствительные глазки могут быть связаны с любой из сетей. Эволюция нервной системы иглокожих настолько специфична, что заслуживает отдельного глубокого рассмотрения. Однако всерьёз считать иглокожих эволюционными
преемниками дорсального и вентрального слияния нервных стволов невозможно (см. рис. II-11, в). Причиной для такого вывода является наличие у иглокожих трёх параллельных нервных систем, хотя у всех видов только одна эктоневральная сеть. По-видимому, она и была первичным нервным образованием в этой группе. Апикальная и гипоневральная сети возникли у иглокожих вторично, что подтверждается
их вариабельностью или полным отсутствием у многих представителей этого типа.
Глава I. Принципы работы и организации мозга
2. Тропизмы и таксисы простейших
3. Функциональная организация нервной системы
4 .Отношение массы мозга и тела.
5. Энергетические расходы нервной систем
6. Потребление мозгом кислорода
7. Гематоэнцефалический барьер.
8. Заряды мембраны нервных клеток
9. Синаптические контакты нервных клеток
10. Типы объединения нервных клеток
11. Нервная система беспозвоночных
12. Нервная система позвоночных
Органы чувств и эффекторные системы
13.Рецепторы и органы чувств
14. Эффекторные системы
Память и забывание
15. Механизмы памяти
16. Морфологические принципы памяти
17. Признаки мышления
18. Биологические проблемы мышления
Глава II. Возникновение нервных клеток и мозга
19.Происхождение нейронов и пронейрональной сети
20. Нервная система с радиальной симметрией
21. Билатеральная нервная система
22. Нервная система членистоногих
23. Нервная система моллюсков
24. Эволюция ганглиозной нервной системы
Нервная система хордовых
25. Теории происхождения хордовых
26. Происхождение нервной системы хордовых
27. Мозг первичноводных позвоночных
28. Возникновение отделов головного мозга
29. Формирование мозга позвоночных
30. Особенности строения нервной системы
31. Проблемы выхода амфибий на сушу
32. Появление конечностей
33. Выход амфибий на сушу
Глава III. Становление мозга амниот
35. Многообразие низших амниот
36. Общий план строения нервной системы рептилий
37. Ассоциативный центр мозга рептилий
38. Условия возникновения мозга рептилий
39. Происхождение неокортекса
40. Адаптивная радиация архаичных рептилий
41. Биологическое разнообразие птиц
42. Морфологические особенности строения птиц
43. Нервная система и органы чувств птиц
44. Условия возникновения мозга птиц
45. Адаптивная радиация птиц
46. Обзор классификации млекопитающих
47. Особенности нервной системы млекопитающих
48. Органы чувств млекопитающих
49. Возникновение мозга млекопитающих
Теория переходных сред
Список литературы